Bot. Bull. Acad. Sin. (1998) 39: 161_165
Lin and Kao Effect of exogenous H2O2 on leaf senescence
Effect of oxidative stress caused by hydrogen peroxide on
senescence of rice leaves
Jaw-Neng Lin and Ching Huei Kao1
Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China
(Received September 25, 1997; Accepted February 26, 1998)
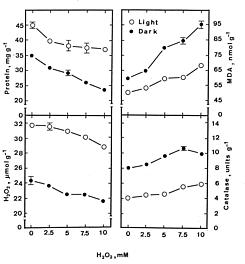
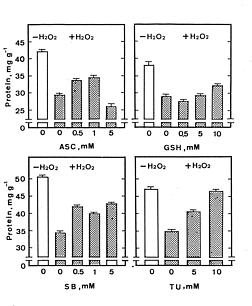
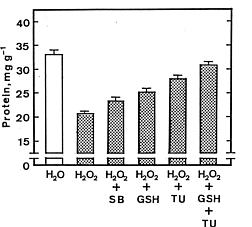
Abstract. Lipid peroxidation is considered to be an important mechanism of leaf senescence. The peroxidation of lipids can be initiated by free radicals. H2O2 itself is active oxygen species and can react with superoxide radicals to form more reactive hydroxyl radicals in the presence of trace amounts of Fe or Cu. Thus, it is of interest to investigate the relationship between lipid peroxidation and H2O2-promoted senescence in detached rice leaves. H2O2 effectively promoted senescence and increased malondialdehyde levels in detached rice leaves under both light and dark conditions. However, the promotion of senescence by the exogenous level of H2O2 is not associated with an increase in its endogenous level. Thiourea (a scavenger of hydroxyl radical) almost completely prevented H2O2-promoted senescence and the H2O2-induced decrease in superoxide dismutase and ascorbate peroxidase activities in light and darkness. It seems that promoted senescence by H2O2 in detached rice leaves is associated free radical-induced lipid peroxidation.
Keywords: Free radical scavengers; H2O2; Leaf senescence; Lipid peroxidation; Oryza sativa; Oxidative stress.
Abbreviations: APOD, ascorbate peroxidase; ASC, ascorbic acid; GR, glutathione reductase; GSH, reduced glutathione; MDA, malondialdehyde; SB, sodium benzoate; SOD, superoxide dismutase; TU, thiourea.
Introduction
Hydrogen peroxide (H2O2) is a constituent of oxidative plant metabolism. It is a product of peroxisomal and chloroplastic oxidative reactions (Del Rio et al., 1992). H2O2 itself is an active oxygen species. H2O2 can also react with superoxide radicals to form more reactive hydroxyl radicals in the presence of trace amounts of Fe or Cu (Thompson et al., 1987). The hydroxyl radicals initiate self-propagating reactions leading to peroxidation of membrane lipids and destruction of proteins (Asada and Takahashi, 1987; Bowler et al., 1992; Halliwell, 1987). It has been reported that H2O2 promotes senescence of detached leaves (Begam and Choudhuri, 1992; Mondal and Choudhuri, 1981; 1982; Parida et al., 1978; Sarkar and Choudhuri, 1981) and induction of senescence is accompanied by an increase in endogenous H2O2 level (Mondal and Choudhuri, 1981). Lipid peroxidation is considered to be an important mechanism of leaf senescence (Dhindsa et al., 1981; 1982; Kunnert and Ederer, 1985; Strother, 1988; Thompson et al., 1987). The peroxidation of lipids can be initiated by free radicals (Girotti, 1985; Thompson et al., 1987). Thus, accelerated senescence in leaves could be the result of an H2O2-mediated increase in rates of oxidative reactions. We thus examined the relationship between lipid peroxidation and H2O2-promoted senescence in detached rice leaves.
Materials and Methods
Rice (Oryza sativa L. cv. Taichung Native 1) was cultured as previously described (Kao, 1980). The apical 3-cm segments excised from the third leaves of 12-day-old seedlings were used. A group of 10 segments was floated in a Petri dish containing 10 mL of test solutions. Incubation was carried out at 27°C in light (40 µmol m-2 s-1) or in darkness.
For protein determination, leaf segments were homogenized in 50 mM sodium phosphate buffer (pH 6.8). The extracts were centrifuged at 17,600 g for 20 min, and the supernatant liquids were used for determination of protein by the method of Bradford (1976). Protein levels were expressed as mg g-1 fresh weight. Malondialdehyde (MDA) was extracted with 5% (w/v) trichloroacetic acid and determined according to Heath and Packer (1968). MDA level is routinely used as an index of lipid peroxidation and was expressed as nmol g-1 fresh weight.
The H2O2 level was colorimetrically measured as described by Jena and Choudhuri (1981). H2O2 was extracted by homogenizing 50 mg leaf tissue with 3 mL of phosphate buffer (50 mM, pH 6.5). The homogenate was centrifuged at 6,000 g for 25 min. To determine H2O2 level, 3 mL of extracted solution was mixed with 1 mL of 0.1% titanium sulphate in 20% H2SO4 (v/v), and the mixture was then centrifuged at 6,000 g for 15 min. The intensity of the yellow colour of the supernatant was measured at 410 nm. H2O2 level was calculated using the extinction coefficient (0.28 µmol-1cm-1).
1Corresponding author. Fax: 886-2-23620879; E-mail: kaoch@cc.ntu.edu.tw