Bot. Bull. Acad. Sin. (1998) 39: 167_174
Chen and Lee Polyamines and hyposaline stress in Ulva fasciata
Hyposaline effect on polyamine accumulation in Ulva fasciata (Ulvales, Chlorophyta)
Tse-Min Lee1 and Ming-Hwa Chen
Institute of Marine Biology, National Sun Yat-sen University, Kaohsiung, Taiwan 80424, Republic of China
(Received November 4, 1997; Accepted March 3, 1998)
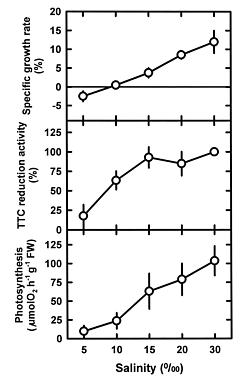
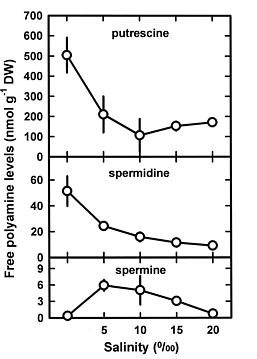
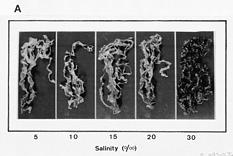
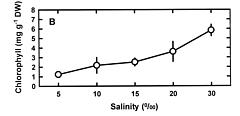
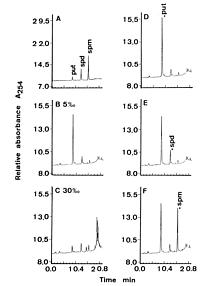
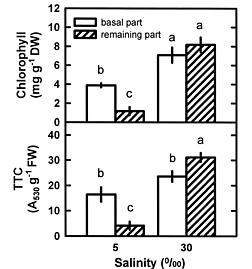
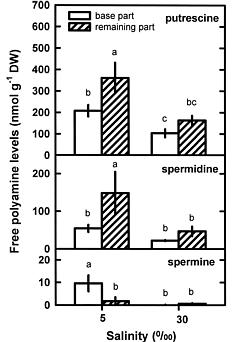
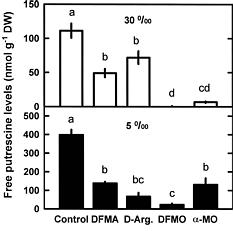
Abstract. This study was conducted to investigate changes of free polyamine levels in response to hyposaline stress in Ulva fasciata Delile. Free putrescine, spermidine, and spermine are present in this alga. As salinity decreased from 30 to 5, specific growth rate (%/d), TTC reduction activity, net photosynthetic O2 evolution rate and chlorophyll levels decreased. Plants grown at 5 bleached except the basal part near rhizoidal portion. Free putrescine maintained at a constant level at salinity over the range of 10~30 and increased three fold following 5 treatment. Free spermidine also increased when salinity fell below 15. In plants grown at 5, free putrescine and spermidine accumulation in the basal part was less than in the remaining part. Free spermine levels increased as salinity decreased from 30 to 10, but dropped sharply at 5. Both a-difluoromethylarginine (0.2 mM) and D-arginine (1 mM), inhibitors of arginine decarboxylase (EC 4.1.1.19), and also both a-difluoromethylornithine (0.2 mM) and a-methylornithine (0.2 mM), inhibitors of ornithine decarboxylase (EC 4.1.1.17), inhibited the 5-induced free putrescine accumulation. Overall, the present results suggest that an extreme hyposaline condition (5) induced a significant accumulation of free putrescine and spermidine in U. fasciata and that ADC and ODC each contributed to free putrescine accumulation. The relationships between accumulated putrescine and hyposaline injury are discussed.
Keywords: Hyposaline stress; Putrescine; Spermidine; Spermine; Ulva fasciata.
Introduction
Polyamines are involved in the regulation of plant growth and development (Evans and Malmberg, 1989; Galston and Kaur-Sawhney, 1987). In plants, putrescine is synthesized from arginine or ornithine through either arginine decarboxylase (ADC; EC 4.1.1.19) or ornithine decarboxylase (ODC; EC 4.1.1.17), and is then converted to spermidine/spermine by the addition of propylamine group from decarboxylated S-adenosylmethionine that is derived from S-adenosylmethionine (SAM) by the action of S-adenosylmethionine decarboxylase (SAMDC; EC 4.1.1.50) (Evans and Malmberg, 1989). A variety of studies have shown that polyamines, especially putrescine, accumulate under stresses, and these accumulated polyamines are closely associated with plants' responses to stress (Evans and Malmberg, 1989). It is suggested that, under stresses, polyamines acts as polycations in maintaining membrane and nucleic acid integrity at cellular pH (Galston and Kaur-Sawhney, 1987).
The effects of salinity stress on polyamine biosynthesis were analyzed in several plant systems (Basu et al., 1988; Krishnamurthy and Bhagwat, 1989; Lin and Kao, 1995; Priebe and Jager, 1978), and the pattern of polyamine metabolism in response to salinity stress seems to be dependent on the plant systems and/or duration of ex
posure to salinity stress. In pea (Anderson and Martin, 1973) and barley (Smith, 1973), neither putrescine nor spermidine accumulate when exposed to increasing salinity. A similar trend was also found in Vicia faba (Priebe and Jager, 1978). In the case of Brassica campestris, exposure to short-term hypersaline stress increased levels of polyamines and activities of both ADC and ODC, while long term exposure exhibited little effect (Das et al., 1995). Krishnamurthy and Bhagwat (1989) showed that hypersaline conditions elicited an accumulation of putrescine in salt-sensitive rice cultivars and a significant accumulation of spermidine and spermine in salt-tolerant ones. Lin and Kao (1995) showed a contrasting result, finding that increasing NaCl levels lead to a decrease in free putrescine levels but an increase in spermidine levels in a salt-sensitive rice cultivar, cv. Taichung Native 1.
It is well documented that polyamines, especially putrescine and spermidine, exist in prokaryotic and eukaryotic algae (Badini et al., 1994; Hamana et al., 1983, 1990; Hamana and Matsuzaki, 1982, 1985). Uncommon polyamines (norspermidine and norspermine) are also widespread in eukaryotic algae (Hamana and Matsuzaki, 1982), playing a vital role in cell growth and development. In both Chlorella vulgaris Beijernck var vulgaris fa vulgaris (Cohen et al., 1984) and Euglena gracilis Z (Villanueva et al., 1980a), a rise of polyamines, especially putrescine derived from ODC, before cell division, is believed to be correlated with DNA replication. Environmental changes also influence the biosynthesis of polyamines in algae.
1Corresponding author. Tel: 886-7-5252000 ext. 5110; Fax: 886-7-5255100; E-mail: tmlee@mail.nsysu.edu.tw