Bot. Bull. Acad. Sin. (1998) 39: 175_179
Su et al. Root sugar level and flooding tolerance
Root sugar level in luffa and bitter melon is not referential to their flooding tolerance
Pai Hsiang Su, Tsui Hui Wu, and Chin-Ho Lin1
Department of Botany, National Chung-Hsing University, Taichung, Taiwan 40227, Republic of China
(Received November 25, 1997; Accepted April 2, 1998)
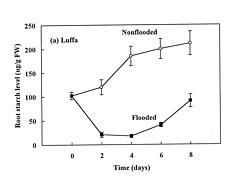
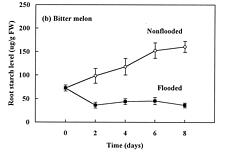
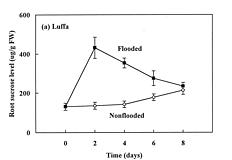
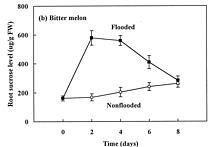
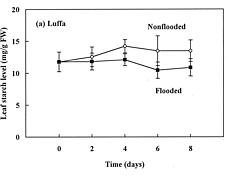
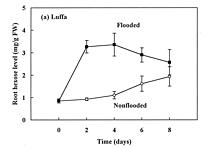
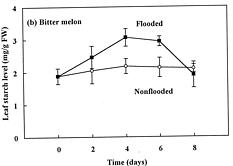
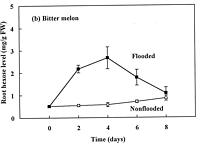
Abstract. The effects of soil flooding for 8 days on root and leaf carbohydrate levels in relation to flooding tolerance in two cucurbitaceous plants: luffa (Luffa cylindrica Roem. cv. Cylinder #2), a flood-tolerant species, and bitter melon (Momordica charantia L. cv. New Known You #3), a flood-intolerant species, were investigated. With flooding, the levels of starch in luffa and bitter melon roots decreased rapidly, indicating that root starch was readily mobilized under flooding conditions. Significant accumulation of leaf starch was observed only in the intolerant bitter melon suggesting reduced phloem transport. Sucrose and hexose (glucose and fructose) levels in flooded roots of both species increased several fold with initial flooding, and were maintained at a higher level than those of non-flooded control during the flooding period. These results implied that starch accumulation in leaves does not result in the deficiency of sugar in flooded roots, and root sugar level is not critical for the flooding tolerance of luffa and bitter melon.
Keywords: Bitter melon; Carbohydrates; Flooding tolerance; Luffa cylindrica; Momordica charantia.
Introduction
The tolerance of a plant to anoxia or soil flooding has been postulated to be determined by the level of sustainable sugar in the root. Saglio et al. (1980) suggested that under anoxia, there is a positive relationship between adenylate energy charge and metabolic activity through the anoxic fermentation pathway, that is, the higher the metabolic activity, the higher the energy charge that will be maintained under anoxia. The anoxic fermentation in roots flows through glycolysis, and the fermentation rate in roots was thus thought to be controlled by the available supplement of soluble sugars. Vartapetian (1991) reported that exogenous glucose postponed the onset of ultrastructural disintegration of excised pea root cells. Zhang and Greenway (1994) found that exogenous sugar prevent depletion of sugar supplies in beetroots, and enhance ethanolic fermentation. Wample and Davis (1983) concluded that the accumulation of starch in leaves of flooded sunflower plants is a reflection of reduced phloem transport. Those studies generally implied that flooding results in the reduction of phloem transport, causing a further accumulation of photosynthate in leaves and a deficiency of carbohydrates in roots. Carbohydrate availability in roots during flooding might be a key factor determining the flood-tolerance of a plant. However, the observation of large accumulation of sucrose in roots of flooded alfalfa, a flood-intolerant species (Barta, 1988; Castonguay et al.,
1993), suggested that the level of sugar in roots might not be a limiting factor to root anaerobic metabolism. Therefore, whether the sugar level in a root determines the tolerance of plant to flooding or not needs further elucidation.
In the present study, the carbohydrate levels in leaves and roots of luffa (Luffa cylindrica Roem. cv. Cylinder #2, a flood-tolerant species) and bitter melon (Momordica charantia L. cv. New Known You #3, a flood-intolerant species, Liao and Lin, 1994) were compared. We intended to analyze the relationship between sugar level in flooded roots of luffa and bitter melon and flood-tolerance.
Materials and Methods
Material Preparation
Seeds of luffa (Luffa cylindrica Roem. cv. Cylinder #2) and bitter melon (Momordica charantia L. cv. New Known You #3) were purchased from Known You Seed Company (Kaohsiung, Taiwan). The soaked seeds were germinated in vermiculite for 7 days, and the germinated seedlings were planted individually in a 1.6 dm3 plastic pot filled with a 2:1:1 (on volume basis) mixture of vermiculite, perlite, and peat. The seedlings were cultivated in a nylon net house with glass roof without temperature control. During the experimental periods, the averaged day/night temperatures were 30/26°C. Approximately 0.3 g per pot of a complete fertilizer (N:P:K=20:20:20), Hyponex No. 2 (Hyponex Co., Inc., Copley, Ohio, USA), was supplied to potted plants every week until the application of the treatments. Plants reaching the twelve-leaf stage (approximately 4 weeks) were used for the experiment.
1Corresponding author. Tel: 886-4-2862511; Fax: 886-4-2862511; E-mail: chlin@mail.nchu.edu.tw