Bot. Bull. Acad. Sin. (1998) 39: 181_186
Chen et al. A pectin lyase from Pythium splendens
Purification and characterization of a pectin lyase from Pythium splendens infected cucumber fruits
Wei-Chen Chen1, Huann-Ju Hsieh1, and Tsung-Che Tseng2,3
1Department of Plant Pathology and Entomology, National Taiwan University, Taipei, Taiwan, Republic of China
2Institute of Botany, Academia Sinica, Nankang, Taipei, Taiwan, Republic of China
(Received February 24, 1998; Accepted April 3, 1998)
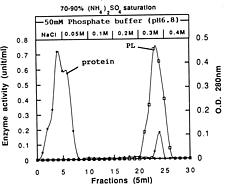
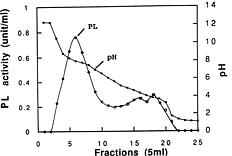
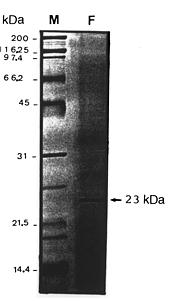
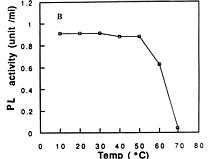
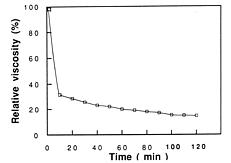
Abstract. A pectin lyase (PL, EC 4.2.2.10) was produced by Pythium splendens Braun in infected cucumber fruits. The enzyme was purified to near homogeneity by ammonium sulfate fractionation, CM - Sepharose column chromatography, and preparative isoelectric focusing. The enzyme had a molecular mass of 23 kDa as determined by SDS-PAGE. The pI of the PL was 8.0, which was also the optimum pH for the enzyme activity. The enzyme was stable at 4_50°C, but beyond 50°C, its activity decreased rapidly. The pectin lyase degraded high methyl-esterified pectin by trans-elimination in an endo-manner, and was stimulated by some divalent cations including Ca2+, Mg2+, and Sr2+. Addition of Zn2+ inhibited the enzyme activity by 36%. The purified PL caused rapid maceration of potato and cucumber discs. This is the first report of a PL produced by Pythium splendens, a plant pathogenic fungus which causes seedling damping-off and fruit rot in cucumber.
Keywords: Cucumber (Cucumis sativa L.); Pectin lyase; Pythium splendens.
Introduction
Plant pathogenic fungi produce an array of extracellular degradative enzymes that may be important in pathogenesis (Walton, 1994). Most attention has focused on enzymes that depolymerize pectin (Collmer and Keen, 1986; Cooper, 1984). Pectin lyase (PL, EC 4.2.2.10) catalyzes the trans-elimination of pectin and has been isolated from a variety of bacterial and fungal pathogens (Collmer and Keen, 1986; Linhardt et al., 1986). Its role in tissue maceration by plant pathogenic bacteria has been demonstrated using mutants that lack PL activity (Beaulieu et al., 1993). Recently, several reports have suggested that PL may be a pathogenicity factor in fungal-plant interactions (Crawford and Kolatlukudy, 1987; Wattad et al., 1994; Wattad et al., 1995; Wijesundera et al., 1984).
Pythium splendens Braun is an economically important soil borne plant pathogenic fungus which causes seedling damping-off and fruit rot in cucumber (Cucumis sativa L.). This fruit rot is apparently caused by the activity of pectolytic and/or cellulolytic enzymes, but this is still unverified. In many pythium diseases, secretion of cell wall-degrading enzymes has been reported (Cherif et al., 1991; Dube and Prabakaran, 1989). The loosening of primary walls by pectolytic enzymes may cause osmotic stress and subsequent cell death (Hall and Wood, 1973). The softening of cucumber fruit in cottony leak disease, caused by Pythium aphanidermatum, has been demostrated to be related to the activities of endo-polygalacturonase and cellulase (Zamski and Peretz, 1996).
However, neither the production of pectolytic enzymes nor cell wall-degrading enzymes are available for P. splendens infected cucumber. This is the first report on the production, purification, and characterization of a pectin lyase for P. splendens infected cucumber fruits.
Materials and Methods
Pythium splendens was isolated from the roots of a diseased cucumber seedling near Taipei and was maintained routinely on V-8 agar slants (200 ml V-8 juice, 3 g CaCO3, 15 g agar in 800 ml distilled water) at 28°C.
Inoculum
Three discs (10 mm dia) were cut from a V-8 agar plate and were grown in a 10 ml V-8 liquid medium in a 50 ml Erlenmeyer flask for 4 days at room temperature. Inocula were obtained after homogenization and centrifugation of the culture at 20,000 g for 10 min. The sporangia-containing pellets were resuspended in distilled water and used as inoculum.
Enzyme Preparation
Cucumber fruits (Cucumis sativa L.) were purchased from a local supermarket. They were surface sterilized by washing in 10% (v/v) sodium hypochlorite for 10 min and then rinsed twice in distilled water. The fruits were aseptically punched with a needle before inoculation with the sporangium suspension (2×103 sporangia/ml). Controls were inoculated with several drops of sterilized water on the fruit surface. Inoculated fruits were kept in chambers under 100% relative humidity at 25°C for 3 days. Rotted
3Corresponding author. Tel: 886-2-27823050 ext. 424; Fax: 886-2-26515042.