Bot. Bull. Acad. Sin. (1998) 39: 191_198
Lin and Shao Temporal changes in intertidal seagrass beds
Temporal changes in the abundance and growth of intertidal Thalassia hemprichii seagrass beds in southern Taiwan
Hsing-Juh Lin1 and Kwang-Tsao Shao
Institute of Zoology, Academia Sinica, Taipei 115, Taiwan, Republic of China
(Received August 26, 1997; Accepted April 9, 1998)
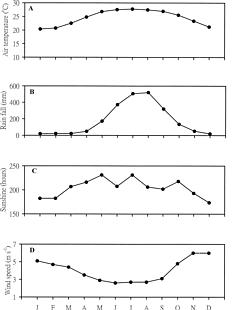
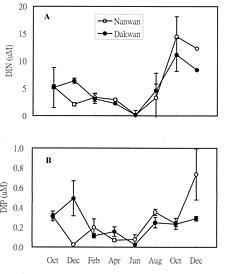
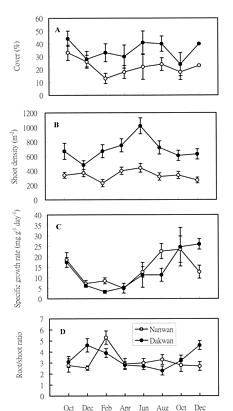
Abstract. A bimonthly study of temporal changes in abundance and growth of the tropical seagrass Thalassia hemprichii on intertidal reef flats was conducted in southern Taiwan from October 1995 to December 1996. Thalassia hemprichii showed a marked temporal pattern in percentage cover, shoot density, and specific growth rate. There were no significant differences in the variables of T. hemprichii between the study sites. The percentage cover showed a unimodal or bimodal pattern in which one peak occurred in June or August and the other occurred in October. The shoot density was highest in June and lowest between December and February. The specific growth rate peaked in October or December, but declined in February or April. However, the temporal patterns in root:shoot ratio of biomass were different. Higher root:shoot ratios were observed in December or February, and lower between April and August. The above-ground biomass was always smaller than the below-ground biomass. Canonical correlation analysis indicated that temperature, water-column nutrient concentration and rainfall were positively correlated, and daylight exposure time and wind speed negatively correlated, with the dynamics of T. hemprichii. Among these variables, wind speed and rainfall were most responsible for the observed temporal changes in southern Taiwan, which has a monsoonal climate and distinct wet and dry seasons.
Keywords: Canonical correlation analysis; Monsoon; Rainfall; Tropical seagrass.
Introduction
Seagrasses are one of the most conspicuous communities of the shallow waters of the world. They comprise only about 50 species worldwide (den Hartog, 1970), but this number is not indicative of their ecological and economic importance. Many investigations have demonstrated that seagrass beds are among the most productive areas of aquatic ecosystems (e.g. Zieman and Wetzel, 1980). They are widely acknowledged to be important in providing food resources (e.g. Klumpp et al., 1989), as well as nursery and feeding grounds, for invertebrates (e.g. Orth et al., 1996) and juvenile fishes (Bell and Pollard, 1989). Furthermore, they may help in stabilizing sediments and preventing coastal erosion and siltation of coral reefs (Fonseca and Fisher, 1986). Nevertheless, a worldwide decline in the abundance of seagrasses due to man-made perturbations has been increasingly reported (e.g. Cambridge and McComb, 1984; Orth and Moore, 1983).
Seagrass communities have long been recognized to be dynamic on a variety of temporal scales (den Hartog, 1970). Wherever man-made perturbations occur, it is necessary to know the extent of the changes that these perturbations may cause (e.g. Lin et al., 1996). For this reason, determining a baseline or control in order to track the ex
tent of man-made changes is essential, and temporal fluctuations in seagrass communities must be documented before any significant perturbation occurs. At present, the available seagrass dynamics data are almost exclusively from temperate regions or the tropical Caribbean (e.g. Zieman, 1975). Only recently have the dynamics of tropical seagrasses along the western Pacific begun to be studied (Brouns, 1985; Erftemeijer and Herman, 1994; Lanyon and Marsh, 1995). These studies, however, have been conducted exclusively in the tropical southern hemisphere between the latitudes of 5° and 15°S. In addition, most of them have been conducted in mixed seagrass beds. Since interspecific interactions are believed to mask seasonal dynamics of seagrasses to some degree (Brouns, 1985; Lanyon and Marsh, 1995), separating abiotic from biotic effects in tropical mixed seagrass beds is difficult (Poiner et al., 1989).
Taiwan lies near the northern latitudinal limit for the geographical distribution of Thalassia hemprichii (Ehrenb.) Aschers., which is one of the two most widely distributed tropical seagrasses in the western Pacific (Mukai, 1993). Prior seagrass studies in Taiwan have all been limited to structural aspects (Doebler, 1984; Huang, 1989; Huang, 1995; Mok et al., 1993) and provide little information on dynamics. The purposes of this study are first, to determine temporal changes in the abundance and growth of T. hemprichii in southern Taiwan, and secondly, to examine which environmental factors are most responsible for the observed temporal changes of T. hemprichii.
1Corresponding author. Tel: +886-2-27899545; Fax: +886-2-27883463; E-mail: hjlin@gate.sinica.edu.tw