Bot. Bull. Acad. Sin. (1998) 39: 199_203
Chang et al. Daylength and flowering in tuberose
Daylength affects protein pattern and flowering in tuberose (Polianthes tuberose L.)
Yu-Pei Chang1, Shih-Fen Ding1, Chin-Chih Chou1, Bo-Shiun Du2 and Wen-Shaw Chen1,3
1Department of Biological Sciences, National Sun Yat-Sen University, Kaohsiung City, Taiwan, Republic of China
2Department of Horticulture, National Chia-Yi Institute of Technology, Chia-Yi City, Taiwan, Republic of China
(Received December 8, 1997; Accepted April 2, 1998)
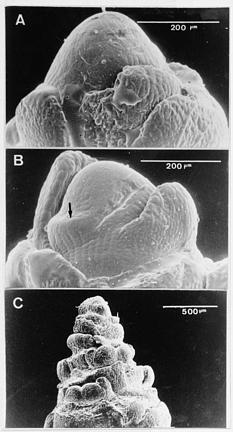
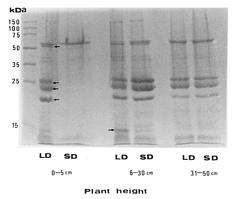
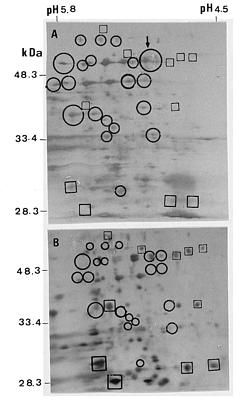
Abstract. Tuberose (Polianthes tuberose L. cv. Double) was exposed to 8 h and 16 h daylengths, to investigate the effects of these conditions on floral initiation and development. Corm apices remained vegetative when plant height was 0_5 cm at 8 h daylength. Floral initiation was attained at five-thirteenth (38%) of this stage at 16 h daylength. However, no significant difference in floral initiation or development of tuberose was found between 8 and 16 h daylengths when plants were over 5 cm in height. When tuberose seedlings were exposed to 16 h daylength, differences in protein pattern of the corm tissues were observed at 19, 23, 27, and 57 kDa. Furthermore, a significant protein band at 11 kDa was found in 16 h daylength but not in 8 h daylength, when plant heights were 6 to 30 cm. However, no significant difference in protein patterns was found between daylengths when plant heights were over 30 cm. The electrophoregrams obtained by two-dimensional electrophoresis allowed characterization of about 185 protein spots in corm tissues when plants were 0_5 cm in height and exposed to 8 and 16 h daylengths. At this stage, 20 newly synthesized proteins were observed in 16 h daylength, and 4 other polypeptides showed significant increase in 16 h, as compared to 8 h, daylength. The most significant peptides were 57 kDa protein with isoelectric point 5.1. These results are discussed in connection to flowering.
Keywords: Daylength; Flowering; Polianthes tuberose; Polypeptide.
Introduction
The effects of photoperiod and temperature on apical meristem development and the transition to reproductive growth have been studied extensively over the past 70 years (Bernier, 1988; Bolling, 1977; Gibby and Salisbury, 1971; Kadman-Zahavi and Peiper, 1987; Lumsden et al., 1982; McMillan, 1974; Salisbury, 1981). However, little study has focused on the correlation between the perception of environmental signals leading to flower induction and biochemical and molecular changes within the plants (Koorneef et al., 1995). Tuberose is an economically important ornamental crop in tropical and subtropical areas. However, how its flowering behavior is affected by photoperiod remains unknown. Visually, flowering in tuberose begins with floral initiation and then accelerates and sustains rapid elongation of the flower stalk. Investigation of the control of floral initiation and development by photoperiod is expected to be useful information. In this paper, we report the effects of different photoperiod conditions on protein synthesis in corm tissues and the flowering response of tuberose.
Materials and Methods
Plant Material
Tuberose (Polianthes tuberosa L. cv. Double) corms about 1.8 × 2.3 cm in diameter were planted in plastic pots (30 cm wide and 20 cm deep) containing vermiculite: domestic peat: sand (3/2/1 by volume). Seedlings were irrigated with tap water weekly and fertilized with a commercial mixture of 15N-7P-14.2K in two irrigations out of four. Corms were grown in chambers, and kept at 8 and 16 h photoperiods (Philips cool-white fluorescent tubes, 120 µE m-2s-1). Temperature was maintained constantly at 30°C.
Investigation of Flower Bud Development
The experiments were conducted using a randomized complete-block design. Thirteen seedlings from each plant height (0 to 5 cm, 6 to 30 cm, and 31 to 50 cm) under SD and LD conditions, respectively, were used for investigating apical development. The experiment was done three times. Each corm apex was dissected, and replicated (Green and Linstead, 1990) in the form a firm, elastic mold. For the entire procedure, the outer scaly leaves were entirely removed, and two polymer mixtures were used. The cements used were: (A) Mirror-wash 3 polymers (two
3Corresponding author. Tel: 886-7-5252000 ext 3612; Fax: 886-7-7251903.