Bot. Bull. Acad. Sin. (1998) 39: 209_212
Zheng and Ho The rediscovery of Phytophthora polygoni
The rediscovery of Phytophthora polygoni Saw.
Xiabo Zheng1 and H.H. Ho2,3
1Plant Pathogen Research Institute, Nanjing Agricultural University, Nanjing 210095, People's Republic of China
2Department of Biology, State University of New York, New Paltz, New York, 12561, USA
(Received January 14, 1998; Accepted March 16, 1998)
Abstract. Phytophthora polygoni Saw. was first described as a new species by Sawada in 1922, causing leaf spot disease of Polygonum japonicum Meisn. in Taiwan. However, there has been no report of the fungus since its first discovery. Recently, a similar disease occurred on Rumex dentatus L. in Nanjing of Jiangsu Province in mainland China and the pathogen was identified as P. polygoni based on the similar biological characteristics.
Keywords: Phytophthora polygoni; Rumex dentatus.
Introduction
Phytophthora polygoni Saw. was first discovered and described on the leaves of Polygonum japonicum Meisn. (Family Polygonaceae) in Taipei and Taichung of Taiwan causing leaf spot disease from the late winter/early spring to early summer (Sawada, 1922). The disease first appeared as small, round yellow green spots which became dull yellow brown with a tint of purple. Subsequently, the lesions enlarged and turned brown and diffuse, resulting in the death of the lower leaves.
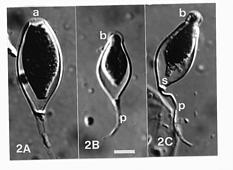
The hyphae of the pathogen were intercellular within the leaf spots, aseptate, colorless and branching with hyphal diameter 5_15 µm (usually 12_14 µm) wide. Appresoria in host cells were mostly obovate in shape. Intercellular hyphae grew through the stomata to form clusters of slender, unbranched or sympodially branched sporangiophores, 15_75 µm long and 1.5_5 µm wide (4_20 sporangiophores per stoma) each terminating in a sporangium with a conical papillum. The sporangia were mostly elongated ellipsoidal to fusiform or pyriform, 17_42 × 4_20 µm. When placed in water, a sporangium germinated either by a germ tube or by releasing motile laterally biflagellated ovoid zoospores with obtuse apex and rounded base, 11_15 × 6_9 µm, encysting to become 10_15 µm in diameter. Neither chlamydospores nor resting sporangia were observed, and no sexual structures were found. Attempts to grow the fungus on culture media failed.
Unfortunately, type specimen of P. polygoni is no longer in existence for re-evaluation, and the fungus has not been reported again since its first description. Efforts were thus made to locate Polygonum and related genera within the family Polygonaceae, with the symptoms as described by Sawada, hoping that P. polygoni could be rediscovered.
Materials and Methods
One genus of the family Polygonaceae, Rumex is a common weed in the Nanjing region of Jiangsu Province in mainland China. Field trips were made to detect and collect Rumex plants whose leaves showed leaf spot disease as described by Sawada (1922).
Freshly collected leaf specimens were prevented from dessication by keeping them in a petri dish with two pieces of moist filter paper on the bottom. They were exposed to natural daylight for 28_48 h at 18_20°C to induce sporangial formation from leaf lesions. The diseased leaf was then examined under microscope at ×150 for the presence of sporangia, which were then carefully scaped off on a glass slide and mounted in a drop of lactophenol, with or without 0.05% cotton blue.
In order to isolate the fungus, two kinds of culture media were used: Lima bean agar (Zhou et al., 1997) and V-8 agar (Ann et al., 1990). Prior to pouring agar medium into sterilized petri dishes, 50 µg/ml each of penicillin, rifampicin, and PCNB (Pentachloronitrobenzene) were added. Diseased leaf was not surface-sterilized, but washed clean with running tap water and allowed to air-dry. Six small pieces of diseased tissue (about 4 × 4 mm) were cut out with a pair of scissors and placed along the edge of an agar plate, at equal distance from one another. At least three agar plates were used for each diseased leaf. In addition, diseased leaf tissues were also surface-sterilized for 5 minutes in 2% chlorine solution prepared by dissolving 1 bleach tablet in 10 ml distilled water. The bleach tablets are produced by Xuhang Chemical Company in Shanghai and marketed by Shanghai Pharmaceutical Company. The sterilized plant material was placed on Lima bean agar and V-8 agar plates without the addition of the above antimicrobial chemicals. The inoculated plates were incubated in dark at 18°C or 25°C for at least one week.
3Corresponding author. Tel: 914-257-3780; Fax: 914-257-3791; E-mail: hoh@npvm.newpaltz.edu