Bot. Bull. Acad. Sin. (1998) 39: 217_223
Luan Immunophilins in animals and higher plants
(Invited review paper)
Immunophilins in animals and higher plants
Sheng Luan
Department of Plant and Microbial Biology, University of California at Berkeley, Berkeley, CA 94720, USA
Abstract. Immunophilins were originally discovered as receptors for a family of immunosuppressive drugs including cyclosporin, FK506, and rapamycin. These drugs suppress immune reaction and are clinically used to prevent graft rejection during organ transplantation. More recently, immunophilins have been found in all eukaryotes ranging from yeast, animals, and higher plants, implicating these proteins in foundamental cellular processes. Biochemical analyses have shown that all immunophilins studied so far possess an enzyme activity involved in protein folding. However, a number of more specific functions for immunophilins have also been discovered in mammalian systems. Although research on plant immunophilins is more primitive, ongoing studies suggest that these proteins may play unique roles in plant growth and development. This article will briefly summarize research activities on both animal and plant immunophilins, a promising new area of current biology.
Keywords: Cyclophilin; FKBP; Protein folding; Signal transduction.
Abbreviations: FKBPs, FK506 and rapamycin-binding proteins; FRAP, FKBP-rapamycin associated protein; RAFT1, Rapamycin and FKBP12 targets.
Contents
Introduction 217
Immunophilins in Animal Systems 218
Targets for Immunosuppression 218
Immunophilins and Protein Folding 218
Immunophilins in Higher Plants 219
An Immunosuppressant-Sensitive Pathway in a Higher Plant 219
Identification of Immunophilins in Higher Plants 219
Summary and Perspectives 220
Acknowledgment 221
Literature Cited 221
Introduction
Intra- and intercellular signal transductions are key processes that regulate cell growth, division and differentiation. The merge of natural product chemistry and cell biology has opened new possibilities for studying intracellular signaling. The logic lies in the fact that many natural products are high affinity ligands to cellular proteins involved in signal transduction.
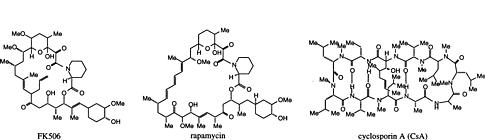
One family of such natural products are immunosuppressive drugs cyclosporin A (CsA), FK506 and rapamycin. As shown in Figure 1, CsA is a cyclic undecapeptide. FK506 and rapamycin, which are structurally related to each other but not to CsA, are macrocyclic lactones (Figure 1). Cellular studies suggest that all three drugs suppress the immune response by blocking
the activation of T lymphocytes (Schreiber, 1991; Kunz and Hall, 1993). CsA and FK506 inhibit a Ca2+-dependent signaling pathway emanating from activation of T cell receptors, and rapamycin blocks a Ca2+-independent pathway required for the proliferation of T cells upon stimulation by lymphokines such as interleukin-2 (Schreiber and Crabtree, 1992; Sigal and Dumont, 1992). In addition to blocking T cell activation, these drugs also have inhibitory effects on signaling pathways in other systems. For example, CsA and FK506 both block the Ca2+-dependent degranulation in mast cells (Hultsch et al., 1991). Rapamycin, on the other hand, has been shown to arrest yeast and some mammalian cells at the G1 phase in the cell cycle (Heitman et al., 1991; Bierer et al., 1990; Dumont et al., 1990; Price et al., 1992). These findings suggest that CsA, FK506 and rapamycin may target molecules that are common signaling components in different systems. What are the target molecules for immunosuppressive drugs?
Tel: (510) 642-6306; Fax: (510) 642-4995; Email: sluan@ nature.berkeley.edu