Bot. Bull. Acad. Sin. (1998) 39: 225_230
Kasai et al. Broccoli and recombinant ACC oxidase
(Invited paper)
Characterization of 1-aminocyclopropane-1-carboxylate (ACC) oxidase in broccoli florets and from Escherichia coli cells
transformed with cDNA of broccoli ACC oxidase
Yuko Kasai1, Hiroshi Hyodo1,3, Yoshinori Ikoma2, and Masamichi Yano2
1Department of Biological Sciences, Faculty of Agriculture, Shizuoka University, Ohya, Shizuoka 422-8529, Japan
2Department of Citriculture, National Institute of Fruit Tree Science, Okitsu, Shimizu 424-0204, Japan
(Received February 9, 1998; Accepted March 24, 1998)
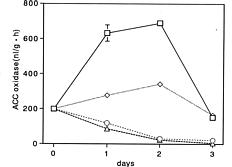
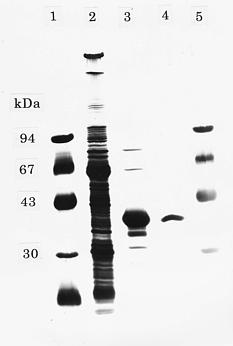
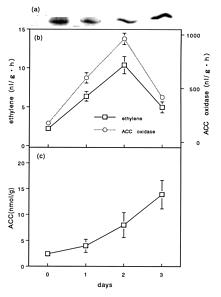
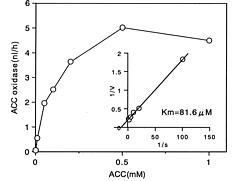
Abstract. During senescence of broccoli (Brassica oleracea L. var. italica) florets (flower buds), the rate of ethylene production increased, almost paralleling the rapid increase in 1-aminocyclopropane-1-carboxylate (ACC) oxidase activity. The increase in ACC oxidase activity was strongly suppressed by the administration of cycloheximide, suggesting that it may have resulted from the de novo synthesis of enzyme protein. ACC oxidase extracted from broccoli florets exhibited the requirements for ferrous ion, ascorbate, and CO2 in its activity besides ACC and O2 as substrates. ACC oxidase was isolated and purified from Escherichia coli cells transformed with cDNA of broccoli ACC oxidase. The recombinant bacterial enzyme was very similar to that of native broccoli enzyme in its enzymatic properties. The molecular mass of the purified enzyme from the transformed cells was estimated to be 37 kDa by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). The polyclonal antibody was raised in a rabbit against the purified enzyme isolated from the transformed bacteria, which was found to cross react with the enzyme of broccoli florets. Western blot analysis showed that ACC oxidase protein was detected at an almost similar level over the progressive stages of senescence.
Keywords: 1-Aminocyclopropane-1-carboxylate (ACC) oxidase; Brassica oleracea; Broccoli florets; Escherichia coli; Senescence; Transformant.
Introduction
Broccoli florets (flower buds) senesce rapidly after harvest at ambient temperatures. Symptoms include a rapid yellowing of florets accompanied by degradation of ascorbic acid and chlorophyll. That endogenous ethylene is involved in the postharvest senescence of florets has been reported (Lieberman and Hardenburg, 1954; Wang, 1977; Aharoni et al., 1985; Makhlouf et al., 1989; Tian et al., 1994). We demonstrated that the rate of ethylene production in broccoli florets increased to a maximum and then declined during senescence, almost paralleling the increasing pattern of ACC oxidase activity (Kasai et al., 1996). It was suggested that the induction of ACC oxidase activity was regulated by endogenous ethylene produced during senescence (Kasai et al., 1996).
In higher plants ethylene is synthesized via methionine-ACC pathway (Yang and Hoffman, 1984; Abeles et al., 1992). In this pathway two crucial enzymes regulate the rate of ethylene synthesis, i.e. ACC synthase and ACC oxidase (Yang and Hoffman, 1984; Abeles et al., 1992; Kende, 1993; Prescott and John, 1996). The enzymology
of ACC oxidase has been studied extensively (Ververidis and John, 1991; Dong et al., 1992; McKeon et al., 1995; Prescott and John, 1996; John, 1997).
We extracted ACC oxidase from broccoli florets and investigated its enzymatic properties (Kasai et al., 1998). In the present study we isolated and purified ACC oxidase from E. coli cells transformed with cDNA of broccoli ACC oxidase, and we compared the enzymatic properties between recombinant bacterial enzyme and native broccoli enzyme. The rabbit antibody has been raised against the purified enzyme from the trnasformed cells, which was used for immunological studies with the broccoli enzyme. We also examined if the increase in ACC oxidase activity in broccoli florets during senescence is derived from de novo synthesis of enzyme protein.
Materials and Methods
Extraction and Assay of ACC Oxidase from Broccoli Florets
Broccoli heads were obtained from a local market in Shizuoka city. Those heads were held at 21±1°C under humidified conditions. Senescence of florets was visualized by the advance of yellowing of florets (Kasai et al.,
3Corresponding author. Tel: 81-54-238-4830; Fax: 81-54-237-3028; E-mail: abhhyou@agr.shizuoka.ac.jp