Bot. Bull. Acad. Sin. (1998) 39: 261_267
Yang and Lin Characteristics from Rhizobium fredii
Cytostructure, lipopolysaccharides, and cell proteins analysis from Rhizobium fredii
Fang-Lin Yang and Liang-Ping Lin1
Graduate Institute of Agricultural Chemistry, National Taiwan University, Taipei 106, Taiwan, Republic of China
(Received January 10, 1997; Accepted June 2, 1998)
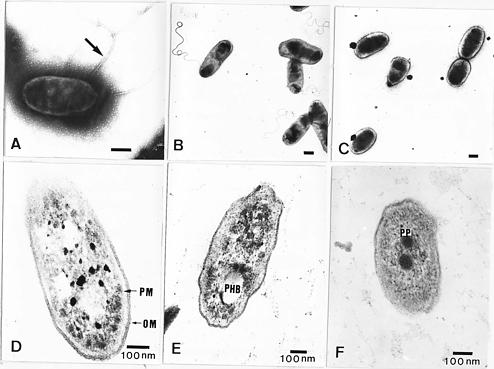
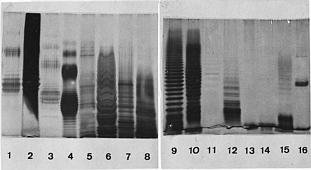
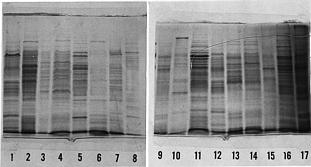
Abstract. The symbiotic roles of rhizobia with legumes have been extensively investigated for agricultural applications. We isolated several strains of rhizobia from Taiwanese soils and analyzed their cellular proteins and lipopolysaccharide (LPS) profiles as well as their ultrastructure by electron microscopy. It was found that the flagella have different forms and locations on the surface of fast-growing and slow-growing rhizobia. In addition, the results of their ultrastructural visualization confirmed that poly-b-hydroxybutyrate (PHB) and polyphosphates are present in rhizobia. The ladder profiles of LPS and cellular proteins on sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE) indicated that different strains of rhizobia possess different profiles, but strains which belong to the same species have similar profiles. It was also shown by Western blot and immunoassay that strain specificity exists between rhizobia cellular proteins and the antisera of anti-rhizobia cells. These results confirm that indigenous R. fredii exhibits a significant difference from other species of Rhizobiaceae.
Keywords: Anti-cellular protein antibody; Anti-LPS antibody; Cell proteins; Cytostructure; Lipopolysaccharides; Rhizobium fredii; Soybean.
Introduction
The taxonomy of rhizobia, based on bacteria-plant cross inoculation groups, was at one time widely used (Elkan, 1992). However, it is now being replaced by numerical taxonomy, serology, metabolic components, and various molecular techniques (Hennecke et al., 1985; Javis et al., 1986; De Lajudie et al., 1994). All of these methods are able to recognize the fast-growing genus Rhizobium and the slow-growing genus Bradyrhizobium by their bacterial growth rate (Jordan, 1984; Elkan, 1992).
Based on our knowledge of Rhizobium-legume symbiosis, the rhizobial cell is involved in interactions with host plants during nodule development (Schmidt et al., 1989; Stacey et al., 1995). The cell surface components of Rhizobium have always been considered important factors in symbiotic effectiveness (Halverson and Stacey, 1986; Hitchcock et al., 1986; Dylan et al., 1990; Gray and Rolfe, 1990; Carlson et al., 1991; Yokota and Sakane, 1991; Niehaus et al., 1993; Ozga et al., 1994) in the specific interactions of rhizobia with legumes. Lipopolysaccharide (LPS) is one component of the bacterial outer membrane and consists of a polysaccharide and lipid A (Hitchcock et al., 1986). Various rhizobial strains differ in LPS composition and serological characters that can nodulate the legume host. Thus, LPS has taxonomic significance in Rhizobiaceae (Yokota and Sakane, 1991). In bacterial taxonomy, the diversity of cellular proteins
in different species is also considered an important factor.
In the present study, we extracted LPS and cellular proteins from several strains of Rhizobium and Bradyrhizobium. These strains were isolated from indigenous soils and purchased from the American Type Culture and Collection (Rockville, MD). The LPS and cellular proteins were analyzed by SDS-polyacrylamide gel electrophoresis and by immunoassay after Western blotting. Based on the results, we can compare the SDS-PAGE profiles of Rhizobium and Bradyrhizobium as well as different species of Rhizobium. In addition, we analyzed the ultrastructural differences between Rhizobium and Bradyrhizobium by electron microscopy, because these results also provide some information for taxonomy, especially in Rhizobiaceae. Significant differences exist in the cellular structure between that of Rhizobium and Bradyrhizobium. From these results, we can obtain more information to confirm the taxonomic significance of indigenous Rhizobium fredii.
Materials and Methods
Strains of Rhizobia and Cultural Conditions
Nine strains of R. fredii and R. loti Lu7 were isolated from Taiwanese soils. The collected legumes (soybean) nodules (ca. 0.3 cm diameter) were washed with distilled water several times. This washing and cleaning procedure was done within 4 h of nodule collection. The nodules were then soaked in 4% H2O2 for 2 min and then washed
1Corresponding author. Fax: (02) 2362-6455.