Bot. Bull. Acad. Sin. (1998) 39: 299_302
Soejima and Peng Aster ageratoides complex in Taiwan
Cytological features of the Aster ageratoides complex (Asteraceae) in Taiwan
Akiko Soejima1 and Ching-I Peng2,3
1College of Integrated Arts and Sciences, Osaka Prefecture University, Osaka 599-8531, Japan
2Institute of Botany, Academia Sinica, Nankang, Taipei 115, Taiwan
(Received July 9, 1998; Accepted September 11, 1998)
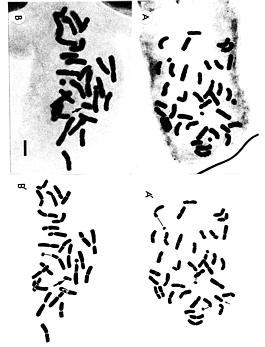
Abstract. Chromosome cytology was examined in two species of the Aster ageratoides complex, A. ageratoides Turcz. (= A. leiophyllus Franch. et Sav.) and A. lasioclada Hayata, in Taiwan. Of the 70 individuals of A. ageratoides sampled, one was found to be diploid (2n = 18) and the rest tetraploid (2n = 36, 37). The occurrence of the diploid A. ageratoides in Taiwan appears to be ancient. Eighteen individuals of A. lasioclada were examined, of which 15 were diploids (2n = 18) and three triploids (2n = 27, 28). The triploid plants were not sympatric with tetraploid plants and could not be distinguished morphologically from the diploids; they may represent spontaneously occurring individuals with one unreduced gamete.
Keywords: Aster ageratoides; Aster lasioclada; Aster leiophyllus; Asteraceae; Chromosome number; Taiwan.
Introduction
Taiwan is a continental island that straddles the Tropic of Cancer. Its location, combined with a tall range of steep, rugged mountains, creates a wide array of environments. Many tropical and subtropical floristic elements are found in the lowlands. In the montane regions of Taiwan, warm to cold temperate elements prevail, which bear a close resemblance to those of mainland China, Japan and the Himalayas (Su, 1992), which together comprise the Sino-Japanese Region of Boreal Kingdom (Good, 1974). Takhtajan (1986) placed Taiwan in the Eastern Asiatic Region of the Boreal Kingdom; Kitamura (1993) placed lowland Taiwan in the Southeastern Asiatic Region and the uplands in the Sino-Japanese Region.
The Aster ageratoides complex [as "A. leiophyllus complex" (Soejima, 1992, 1993); see Ito and Soejima (1995) for taxonomic treatment] is one of the Sino-Japanese elements widely distributed in East Asia. It consists mainly of perennial herbs with elongate stolons; lanceolate, oblong-lanceolate or ovate leaves with three distinct nerves; basal leaves that wither at anthesis; few to many capitula 1_2 cm across arranged in corymbs or loose corymbose panicles; hemispherical involucres; and ray florets with white or pale purple corollas.
Within this complex, a polyploid series ranging from diploid to nanoploid is known (Huziwara, 1957; Peng and Hsu, 1978; Matsuda and Suyama, 1980; Matsuda and
Shinohara, 1985; Irifune et al., 1985; Irifune, 1990; Soejima, 1992, 1993), and the presence of this polyploid series has been a source of confusion in the taxonomy of this complex. In order to clarify the taxonomy and to understand the biological background of this complex, cytological investigation accompanied by morphological studies throughout the range of the A. ageratoides complex is needed. At its southern distribution range in Taiwan, this complex is represented by two species, A. ageratoides Turcz. and A. lasioclada Hayata. Aster morrisonensis can be considered as a derivative of the A. ageratoides complex that has adapted to alpine habitats in Taiwan. Very little is known of the chromosome cytology of this complex in Taiwan, except for a tetraploid count of 2n = 36 from a single plant of A. ageratoides (as "Aster trinervius D. Don ssp. ageratoides (Turcz.) Grierson") reported by Peng and Hsu (1978) without discussion. This study examines the chromosome cytology and morphological features of members of this complex in Taiwan, in an attempt to supplement and compare data, contributing to a better overall understanding of cytological-morphological relationships among all the members of A. ageratoides complex in East Asia.
Materials and Methods
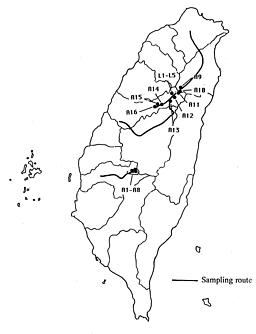
Ninety-two plants were collected from 16 populations of A. ageratoides and 5 populations of A. lasioclada occurring in areas between 865 and 2,200 m altitude in Taiwan (Table 1, Figure 1). The aerial parts of these plants were kept as herbarium voucher specimens (at HAST) for morphological study. The stolons were cultivated at Osaka Prefecture University for cytological investigations. The cytological method follows Soejima (1992).
3Corresponding author. Fax: 886-2-2782-7954; E-mail: bopeng@ gate.sinica.edu.tw