Bot. Bull. Acad. Sin. (1999) 40: 65_71
Wen and Hsiao Genetic differentiation of Lilium longiflorum var. scabrum
Genetic differentiation of Lilium longiflorum Thunb.
var. scabrum Masam. (Liliaceae) in Taiwan using
Random Amplified Polymorphic DNA and
morphological characters
Chia-Szu Wen and Ju-Ying Hsiao1
Department of Botany, National Chung Hsing University, Taichung, Taiwan, Republic of China
(Received September 26, 1997; Accepted July 24, 1998)
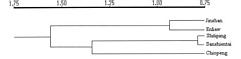
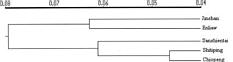
Abstract. Lilium longiflorum Thunb. var. scabrum Masam. is distributed along the northern, eastern, and southern coasts of Taiwan and in some of the outlying islets. It exhibits large amounts of morphological variation among populations in different habitats. Five populations were examined in the present study. In addition to a morphological study, RAPD were used to determine whether the observed morphological variation has a genetic basis and to investigate the variation pattern from different latitudes and different geographical locations. The results of the morphological analyses gave some indication of clinal trends; however, no conclusions can be made because of the limited number of populations studied. In the RAPD study, 140 primers were screened, 9 of which were selected to analyze in all of the samples. The results revealed that, with the exception of the islet Sansientai population, the closer the geographical locations of populations, the closer were their genetic relationships. The exception of islet Sansientai may result from the interruption of gene flow and the effect of genetic drift due to the small size of this islet's population. AMOVA analysis on RAPD data revealed that, of the total variation in the species, 14.08% was attributable to population differences and 85.92% to individual differences within populations when all populations were treated as belonging to a single region. The among population variance component was shown to be highly significant (P<0.001). When two northern populations were treated as a region and the other populations as another region, the result of AMOVA showed that the percentages of variation attributable to the differences between regions, among populations within regions, and among individuals within populations were 5.94% (p<0.001), 10.18% (p<0.001), and 83.88% (p<0.001), respectively.
Keywords: Genetic differentiation; Liliaceae; Lilium longiflorum Thunb. var. scabrum Masam.; RAPD.
Introduction
Lilium longiflorum Thunb. var. scabrum Masam. is a perennial herbaceous monocot of the family Liliaceae. It is distributed along the northern, eastern, and southern coast of Taiwan as well as in some of the outlying islets. Taiwan is on the southern-most boundary of L. longiflorum distribution (Shii, 1983). It possesses large flowers, ranging in color from pure white to white with purple nerves, and has potential commercial value. The species is adapted to diverse habitats and exhibits a high degree of morphological variation. In exposed seashore environments the species may form reef stands about 20 cm in height, while in shaded areas, such as under Pandanus odoratissimus L. f. forest habitat, it can reach a height of 150 cm. Shii (1983) suggested that the distribution of plant height is associated with genetic attributes, local conditions, and plant age. Lilium longiflorum var. scabrum also exhibits changes in leaf width, leaf length, flower shape, and flower color in
different habitats. Wilson (1925) stated that the Taiwanese L. longiflorum was a variety whose name should be L. longiflorum Thunb. var. insular Hort. Masamune (1954) considered it a new variety and named it L. longiflorum Thunb. var. scabrum Masam. Liu & Ying (1978) in the first edition of the Flora of Taiwan adapted the treatment of Masamune. Cheng (1990) studied the variation in twelve populations of the species in Taiwan, employing morphological characters and using Gower's similarity coefficient. The result of cluster analysis indicated that there were three phenomes. Phenomes B and C represented the Pa-Li and Lan-Yu populations, respectively. The other populations belonged to phenome A. He suggested that the observed variation was the result of genetic and environmental variations.
Plant populations may show morphological variations as adaptation to different selection pressure (Nevo et al., 1986; Morrison and Weston, 1985; Hageman and Fahselt, 1990). The adaptation may result from phenotypic plasticity and/or genetic differentiation due to natural selection and other evolutionary forces. Traditional morphological observation alone can not determine the roles of phenotypic plasticity and genetic differentiation on population
1Corresponding Author. Tel: 886-4-2840417~315; Fax: 886-4-2874740.