Bot. Bull. Acad. Sin. (1999) 40: 73_78
Song and Lin Transition of Rhizobium lipopolysaccharides by root exudation
The transition of Rhizobium fredii lipopolysaccharides induced by soybean root exudation
Shuh-Chyung Song and Liang-Ping Lin1
Graduate Institute of Agricultural Chemistry, National Taiwan University, Taipei 106, Taiwan, Republic of China
(Received April 10, 1997; Accepted June 2, 1998)
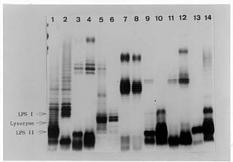
Abstract. The chemical compounds contained in leguminous plant root exudation induce structural and compositional transitions of the lipopolysaccharides of Rhizobium that occur during rhizobia-legume symbiotic infection processes. The root exudate was extracted from the seed germinated solution and seedling roots of soybean. We added the exudation to bacteria cultured medium. Rhizobium and Bradyrhizobium strains were purified through gel filtration chromatography followed by SDS-PAGE analysis. Different chromatographic and SDS-PAGE patterns among the different rhizobia strains were obtained. We determined by means of the gel filtration chromatography that soybean root exudations induced a molecular weight range change in the lipopolysaccharides of these strains. The band of this LPS-II region exhibited diffused patterns in SDS-PAGE as a result of soybean root secretion. The LPS-I region of four strains cultured in TY medium with root exudates exhibited clearer and denser bands than those of rhizobia from TY medium without added soybean root exudates. Based on the composition analysis, we found that the KDO ratio increased in R. fredii TU6, R. meliloti ATCC 9930, R. leguminosarum bv. viciae ATCC 10004 and B. japonicum USDA 110, but decreased in R. fredii KR23, R. loti Lu7, and R. loti ATCC 33669 when soybean root exudation was added to the medium. The hexouronate ratio decreased except in R. loti Lu7. This result was consistent with the SDS-PAGE profile. The LPS-I region of R. loti LU7 showed little variation. It was therefore ascertained that soybean root secretion induces a variation in the structure of rhizobial lipopolysaccharides.
Keywords: Bradyrhizobium japonicum; Lipopolysaccharide; Rhizobium fredii; Root exudation.
Abbreviations: LPS, lipopolysaccharide; Hex, hexose; HexA, hexuronic acid or hexuronate; KDO, 2-keto-3-deoxyoctanate; O-Ag, O-antigen; MWCO, molecular weight cut off; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis.
Introduction
Lipopolysaccharide (LPS) is a component of the outer membrane of Gram negative bacteria and is composed of O-antigen (O-Ag), core oligosaccahride, and lipid A. In the nitrogen fixing bacteria Rhizobium, it has been suspected of playing a role in early symbiotic processes. It has been pointed out in some research results that LPS can affect the ability of rhizobia to nodulate leguminous hosts. For instance, an LPS mutant of Bradyrhizobium japonicum, defective in O-Ag, cannot infect soybean root hair (Puvanesarajah et al., 1987). LPS mutants of R. phaseoli also form a defective infectious thread, so that rhizobial cells cannot be released and form nodules (Cava et al., 1989). Thus, rhizobial LPS is known to be an essential factor in nodule forming processes.
Leguminous root exudations were previously thought to be an important factor in the infection processes and to contain a variety of different molecular weight fractions. Recently, some reports have showed that flavonoid chemicals, secreted in root exudates, can activate nod
genes and regulate the nod gene product as part of a signal compound participating rhizobia-host recognition reaction (Peter et al., 1986; Peter and Long, 1988; Lerouge et al., 1990). The signal compound of R. meliloti was identified as a tetrasaccharide which could be derived from cell wall (Lerouge et al., 1990). In rhizobial extracellular polysaccharides, some alterations in composition, structure, and molecular weight of exopolysaccharides were induced by 1 µM genistein (Dunn et al., 1992). No one has yet reported on the effect on LPS by low molecular weight root exudates although LPS plays an important role in infectious processes.
In this study, we decided to investigate the effect of a mixture, instead of any single chemical substance, because the low molecular weight fraction was always a mixture in the rhizosphere. According to the results of Gaworzeska and Carlile (1982), the low molecular fractions are secreted during 0_3 days of germinating seedling. We therefore collected the root secretion of 3-day-old soybean seedlings for our research. In this paper, we will use the results of gel filtration chromatography and SDS-PAGE to demonstrate that the transition of LPS can be induced by those small molecules.
1Corresponding author: Tel: 886-2-363-5523; Fax: 886-2-362-6455.