Bot. Bull. Acad. Sin. (1999) 49: 93_100
Lee et al. Seasonal acclimation in Gracilaria tenuistipitata
Differences in physiological responses between winter and
summer Gracilaria tenuistipitata (Gigartinales, Rhodophyta)
to varying temperature
Tse-Min Lee1,3, Yuan-Chun Chang1, and Yaw-Huei Lin2
1Institute of Marine Biology, National Sun Yat-Sen University, Kaohsiung, Taiwan 80424, Republic of China
2Institute of Botany, Academia Sinica, Taipei, Taiwan 11529, Republic of China
(Received November 4, 1997; Accepted April 22, 1998)
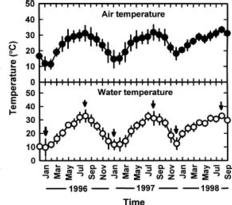
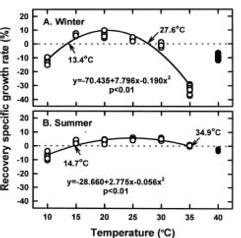
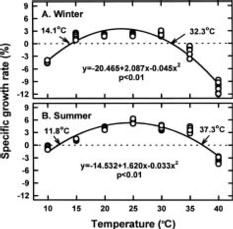
Abstract. Gracilaria tenuistipitata var. liui Zhang et Xia, which is farmed in brackish water ponds of southern Taiwan as a major food of sea abalone, produces poor yields in winter. This study investigated the effects of year, location (pond), and season on the physiological responses of G. tenuistipitata to varying temperature. Neither year nor pond affected the physiological parameters. Season significantly affected the specific growth rate, the recovery specific growth rate, and the levels of free proline. The specific growth rate was different between winter (January in 1996, 1997 and 1998) and summer (August in 1996, 1997 and 1998) plants, which grew in a range of 14.1_32.3°C and 11.8_37.3°C with the maximum at 23 and 24.5°C, respectively. The recovery specific growth rate after transferal to 25°C showed a similar trend. Free proline accumulated only in summer plants at both 20 and 35°C. Thus, the growth and free proline levels at varying temperature are season-dependent events in G. tenuistipitata.
Keywords: Gracilaria tenuistipitata; Growth; Proline; Season.
Introduction
Species of the red algal genus Gracilaria, typically found on the intertidal regions of tropical and subtropical areas, have been studied worldwide because of their increasing market value as agarophytes (Armisen, 1995). Their growth depends mainly on temperature (Gressner, 1970; Hurtado-Ponce and Umezaki, 1987), light (Beer and Israel, 1983), and salinity (Yu and Pedersen, 1990). In Taiwan, several G. species are found (Yang and Chiang, 1982), and the species G. tenuistipitata var. liue Zhang et Xia is now extensively farmed in the brackish water ponds mostly located in Anping, Tainan and is now used as a food source for the cultivation of sea abalone, Haliotis diversiolor supertexta Lischke (Chiang, 1981). The production of G. tenuistipitata is poorer in winter than in summer, possibly due to seasonal changes and overgrowing of competing seaweeds, mainly Enteromorpha and Chaetomorpha (Chiang, 1981).
Macroalgae in many habitats have various physiological mechanisms for responding to environmental changes, and the ability to tolerate environmental disturbances often contributes to their success in marine communities. In addition to genetic adaptation (Kuebler et al., 1991), the survival of a certain species is often determined by its ability to acclimate to environmental changes. Acclimation is known to be a widespread phenomenon in nature, and long-term responses can be observed in the course of a
season. A variety of reports show the occurrence of seasonal changes in the photosynthesis and respiration of intertidal and subtidal algae (Duarte and Ferreira, 1995; Kuppers and Weidner, 1980; Newell and Pye, 1968). It is widely accepted that acclimation allows seaweed to optimize photosynthetic ability and respiration as well, and hence growth, in response to seasonal changes in water temperature (Davison, 1987; Davison and Davison, 1987; Egan et al., 1989). As reported by Friedlander et al. (1987), G. conferta in Israel shows a growth reduction in winter (December-March) and a peak in summer (August), mainly due to extreme seawater temperatures during these seasons: 13°C and 31°C, respectively.
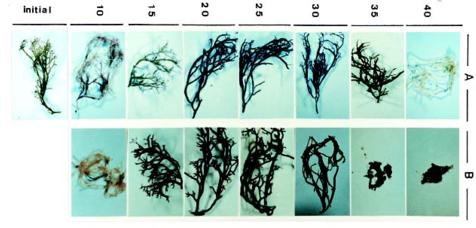
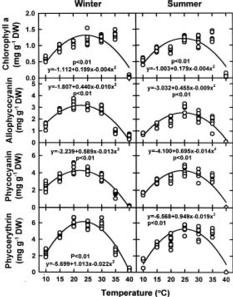
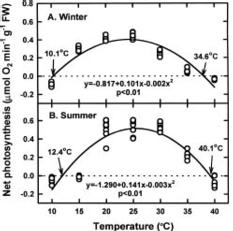
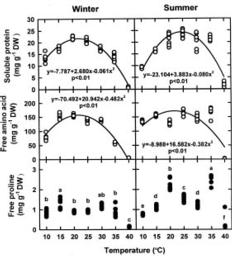
This study was conducted to compare physiological differences between winter- (January in 1996 and 1997) and summer-grown (August in 1996 and 1997) G. tenuistipitata by determining their responses to various temperatures ranging from 10_40°C in laboratory for 5 days. This work was carried out in 1996, 1997, and 1998. The quantitative changes in physiological parameters were examined, including specific growth rate, recovery specific growth rate (i.e. repairing ability), net photosynthesis, pigments (chlorophyll a and phycobiliprotein), and soluble nitrogen compounds (soluble protein and free amino acid). Free proline in algae accumulates under several stresses such as salinity (Bartels and Nelson, 1994; Csonka, 1989) and the presence of heavy metals (Wu et al., 1995a,b). The effects of seasonal acclimation on levels of free proline in response to varying temperature were also determined in G. tenuistipitata.
3Corresponding author. Tel: 886-7-5252000 ext. 5110; Fax: 886-7-5255100; E-mail: tmlee@mail.nsysu.edu.tw