Lu et al. In vivo expression of bifunctional ACSO fusion enzyme
Bot. Bull. Acad. Sin. (1999) 40: 107-114
(Invited Paper)
Expression of a novel ethylene-producing bifunctional fusion enzyme in yeast
Bing Wen Lu, Bing Yu, and Ning Li1
Department of Biology, The Hong Kong University of Science and Technology, Clear Water Bay, Kowloon, Hong Kong
(Received April 28, 1998; Accepted May 19, 1998)
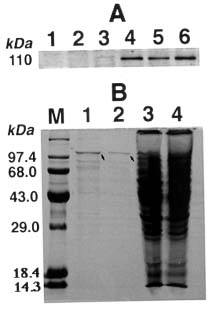
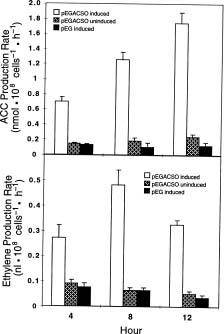
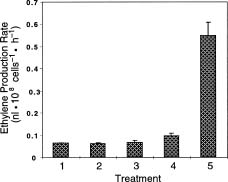
Abstract. The 1-aminocyclopropane-1-carboxylic acid (ACC) dependent ethylene biosynthetic pathway of higher plants was reconstituted in yeast (Saccharomyces cerevisiae). The ACC-dependent ethylene biosynthesis in yeast was catalyzed by a novel bifunctional ACC synthase-ACC oxidase (ACSO) fusion enzyme (Ning Li, Xiang Ning Jiang, Guo Ping Cai and Shang Fa Yang [1996] The Journal of Biological Chemistry 271: 25738_25741). This fusion enzyme ACSO which was further fused to glutathione S-transferase is capable of converting yeast endogenous S-adenosyl-L-methionine (AdoMet) to ethylene. The molecular weight of the fusion enzyme, GST-ACSO, expressed in yeast, as determined by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), was 110 kDa. The ethylene production rate of the yeast cell containing GST-ACSO fusion enzyme was determined to be 21.4 pmol hr-1 per 108 cells at 8 h of galactose induction. The level of ACC, produced as an intermediate during the sequential reactions from AdoMet to ethylene, was found to increase gradually after galactose induction. Because ACSO is capable of producing ethylene from the ubiquitous and prevalent AdoMet in the living eukaryotic cell and the method commonly used to measure ethylene is simple, fast, and extremely sensitive (0.03 parts per billion), we anticipate this bifunctional fusion enzyme to be useful in the near future for research in molecular biology, developmental biology, fermentation, and genetic engineering.
Keywords: ACC oxidase; ACC synthase; ACSO; Bifunctional; Ethylene; Fusion enzyme; Metabolic engineering; Reporter gene; Yeast.
Abbreviations: aa, amino acid; ACC, 1-aminocyclopropane-1-carboxylic acid; AdoMet, S-adenosyl-L-methionine; PCR, polymerase chain reaction; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis.
Introduction
The simple organic molecule, ethylene, has a number of complex biological functions. It regulates the growth and development of higher plants in many ways. These effects of ethylene on higher plants include promoting fruit ripening, flower and leaf senescence, organ abscissions, inhibiting seedling elongation, stimulating root initiation, promoting initiation of the flowering of pineapples, and increasing latex flow in rubber trees (Abeles et al., 1992; Lieberman, 1979; Yang and Hoffman, 1984). Some of ethylene's effects are less desirable, such as when the objective is to prolong the shelf-life of climacteric fruit and flowers. Others are more desirable, such as when the objectives are to enhance the controlled abscission of flowers or young fruits (referred to as thinning), to increase cotton ball opening, to sustain latex flow, to stimulate adventitious root initiation and to promote the female flower differentiation in Cucurbitaceae and the flowering of pineapple plants. To manipulate the extent of the ethylene effect on these agricultural and horticultural crops, many registered
chemical compounds such as aminoethoxyvinyl glycine (AVG) and silver thiosulfate (STS, Ag(S2O3)2_3 ), inhibitors of ethylene biosynthesis and action, respectively, and ethephon, the "liquid ethylene," have been developed (Maynard and Swan, 1963; Owens et al., 1971). Exogenous application of these chemicals onto crops of commercial value has been shown to increase their productivity and quality (Abeles et al., 1992). However, elucidation of the ethylene biosynthesis pathway in higher plants (Adams and Yang, 1979) and recent successful cloning of the key genes involved in the ethylene biosynthesis offer a good opportunity to either enhance or repress ethylene production endogenously via genetic engineering (Hamilton et al., 1990; Nakajima et al., 1990; Sato and Theologis, 1989; Van der Straeten et al., 1990).
Ethylene is synthesized through the "ACC-dependent" pathway in higher plants. In this pathway, the precursor, S-adenosyl-L-methionine (AdoMet), a ubiquitous and prevalent compound involved in many biological reactions including polyamine biosynthesis and biological methylation of polysaccharides, polypeptides, and nucleic acids (Tabor and Tabor, 1984), is first converted to 1-aminocyclopropane-1-carboxylic acid (ACC) and finally to ethylene (Adams and Yang, 1979). Continued production of ethylene via this pathway is sustained by replen
1Corresponding author. Fax: 852-2358-1559; E-mail: boningli@ust.hk