Bot. Bull. Acad. Sin. (1999) 40: 127-134
Kuo et al. Arginine residue in V-ATPase
An essential arginine residue in vacuolar H+-ATPase purified from etiolated mung bean seedlings
Soong Yu Kuo1, Chi Meng Tzeng1, Wen Jye Lin2, Shih Sheng Jiang1, Shu Hsien Hung1, and
Rong Long Pan1,3
1Institute of Radiation Biology, Department of Life Science, College of Life Science, National Tsing Hua University, Hsin Chu, 300 Taiwan, Republic of China
2Department of Radiology, Yuanpei Institute of Medical Technology, Hsin Chu, 300 Taiwan, Republic of China
(Received November 6, 1997; Accepted August 10, 1998)
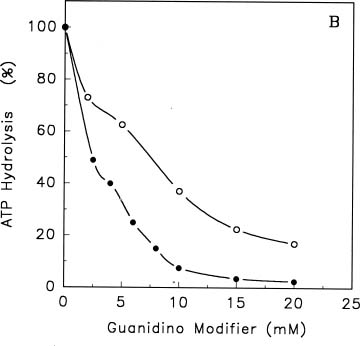
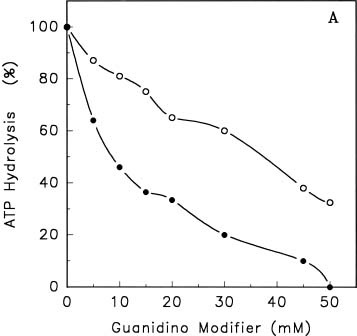
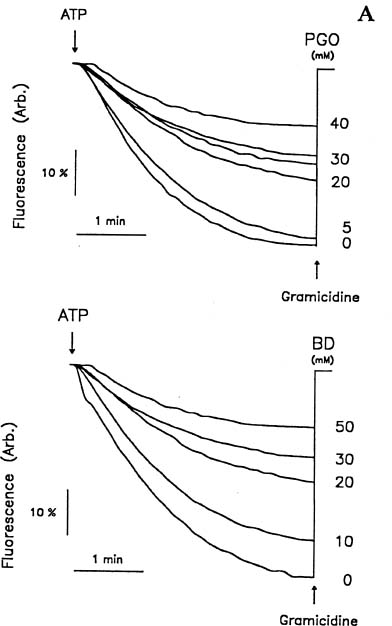
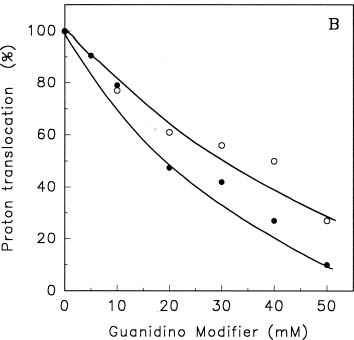
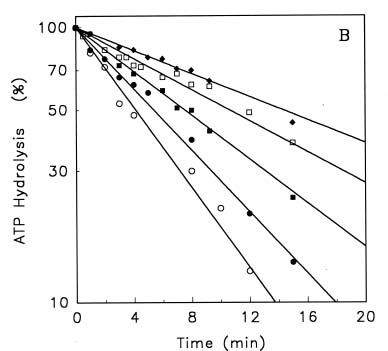
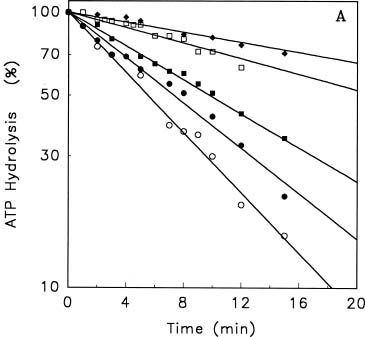
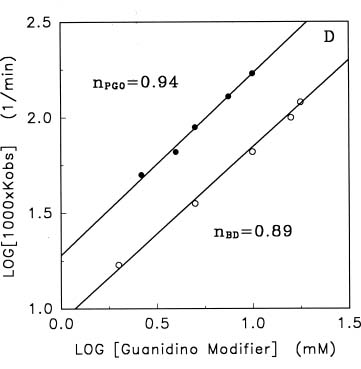
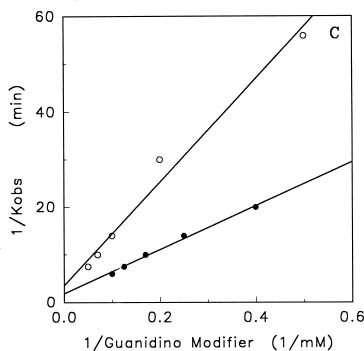
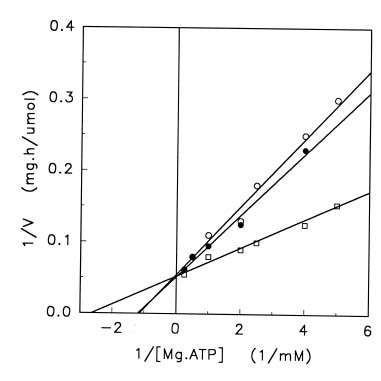
Abstract. Treatments of the tonoplast ATPase purified from mung bean seedlings (Vigna radiata L.) with guanidino modifiers, phenylglyoxal and 2,3-butanedione, caused a marked loss of the ATP hydrolysis activity and proton translocation in a concentration-dependent manner. Kinetic analysis yielded first order rate constants, k2, of 0.416 and 0.227 s-1 and steady-state dissociation constants, Ki, of 19.3 and 24.2 mM for phenylglyoxal- and butanedione-inhibition of vacuolar H+-ATPase, respectively. The reaction order of phenylglyoxal- and butanedione-inhibition was calculated to be 0.94 and 0.89, respectively, suggesting that at least one arginine residue of vacuolar H+-ATPase was modified by both reagents. Lineweaver-Burk plots showed that the mode of inhibition of vacuolar H+-ATPase by both modifiers is competitive. Mg-ATP, the physiological substrate of vacuolar H+-ATPase, but not its analogs, exerted preferentially partial protection against phenylglyoxal and butanedione, indicating that the arginine residue involved in the inhibition of enzymatic activity may be located at or near the active site and directly participate in the binding of the substrate.
Keywords: Chemical modification; H+-ATPase; Mung bean; Proton translocation; Tonoplast.
Abbreviations: BD, 2,3-butanedione; BZ-ATP, 3-O-(4-benzoyl) benzoyladenosine 5'-triphosphate; DCCD, N,N'-dicyclohexyl carbodiimide; DIDS, 4,4'-diisothiocyanostilbene-2,2'disulfonate; NBD-Cl, 7-chloro-4-nitrobenzo-2-oxa-1,3-diazole; NEM, N-ethylmaleimide; PGO, phenylglyoxal.
Introduction
Tonoplast H+-ATPase of higher plants plays a vital role in the secondary active transport of sugars, ions, and metabolites (Sze, 1985; Sze et al., 1992). Tonoplast H+-ATPase is discretely stimulated by Cl- and inhibited by NO3-, DIDS2, NEM, NBD-Cl, and DCCD (Sze, 1985; Sze et al., 1992). Molecular mass of tonoplast H+-ATPase from higher plants was estimated to be 400_600 kDa (Parry et al., 1989; Matsuura-Endo et al., 1990). Vacuolar H+-ATPase contains seven to nine polypeptides upon SDS-PAGE with two major bands of molecular masses of 65_80 kDa (A subunit) and 55_65 kDa (B subunit), and a 16-kDa protein (DCCD-binding proteolipid). The A subunit of vacuolar H+-ATPase is implicated as the catalytic site (Randall and Sze, 1987; Bowman et al., 1986), while the B subunit probably plays a regulatory role (Bowman et al., 1986; Manolson et al., 1985). The DCCD-binding proteolipid subunit might be involved in the proton translocation and is preferentially labeled by radioactive DCCD (Mandala and Taiz, 1986; Manolson et al., 1985; Randall and Sze, 1987).
The cDNA sequences and consequently their deduced amino acid sequences of A subunit from tonoplast H+-ATPases of carrot roots (Zimniak et al., 1988), mung bean (Chiu et al, 1995), cotton (Wilkins, 1993), and Neurospora crassa (Bowman et al., 1988) were recently obtained. Several essential amino acid residues involved in the catalytic activity of vacuolar H+-ATPases from coated vesicles and yeasts were investigated by substrate analogs (Vasilyeva and Forgac, 1996; Zhang et al., 1995; Feng and Forgac, 1994; Manolson et al., 1985; Mandala and Taiz, 1986) and site-directed mutagenesis (Liu et al., 1997; Liu et al., 1996). For instance, the sensitivities of vacuolar H+-ATPase to NEM and NBD-Cl imply the possible presence of cysteine and/or tyrosine residues at the active site (Manolson et al., 1985; Mandala and Taiz, 1986). Further mutation analysis reveals the roles of glycine and glutamate residues in the stability and the catalytic activity of vacuolar H+-ATPase (Liu et al., 1997). Our recent work also indicates, using the substrate analog dialdehyde derivative of ATP, the A subunit of vacuolar H+-ATPase contains a lysine residue essential to the enzymatic activity (Chow et al., 1992; Tzeng et al., 1992). In spite of these results, further efforts are required to identify other amino acid residues significant to the activity of vacuolar H+-ATPase and to elucidate their roles in the enzymatic mechanism.
3Corresponding author. Tel/Fax: (886)-(3)-5742688; E-mail: rlpan@RB.nthu.edu.tw