Bot. Bull. Acad. Sin. (1999) 40: 141-145
Siddique et al. Drought stress effects on photosynthetic rate and leaf gas exchange of wheat
Drought stress effects on photosynthetic rate and leaf gas exchange of wheat
M.R.B. Siddique1, A. Hamid, and M.S. Islam2
Institute of Postgraduate Studies in Agriculture, Gazipur-1703, Bangladesh
(Received June 29, 1998; Accepted September 8, 1998)
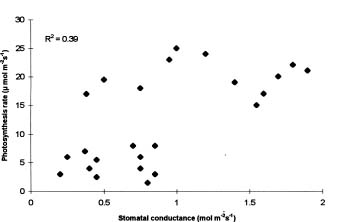
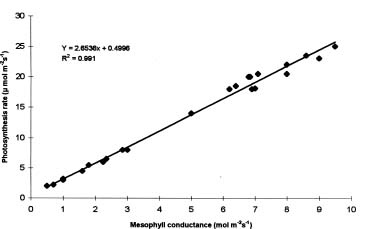
Abstract. Drought stress effects on photosynthetic rate and leaf gas exchange characteristics of four wheat cultivars were evaluated under semi-controlled conditions. Four cultivarsKanchan, Sonalika, Kalyansona, and C306, grown in potswere subjected to four levels of water stress. Among the cultivars, Kalyansona showed the highest photosynthesis rates both at vegetative and at anthesis. Exposure of plants to drought stress led to noticeable decrease in photosynthesis rate, stomatal conductance and mesophyll conductance and a concomitant increase in intercellular CO2 concentration. Plants subjected to drought at the early vegetative stage displayed similar physiological characters subsequently under well-watered conditions as compared with control. Photosynthesis rates decreased with decrease in stomatal conductance, but a weak relationship between them implied that non-stomatal limitations to photosynthesis might have been in operation.
Keywords: Drought stress; Leaf gas exchange; Photosynthesis; Triticum aestivum L.
Abbreviations: Ci, intercellular CO2 concentration; gm, mesophyll conductance; gs, stomatal conductance; Pn, photosynthetic rate; Ta, air temperature; Tl, leaf temperature.
Introduction
Wheat (Triticum aestivum L.) is an important cereal crop that ranks first globally and second in Bangladesh. In Bangladesh wheat is grown generally after the harvest of aman rice on residual soil moisture. Even with substantial improvement of irrigation facilities, 60_65% of the total wheat area remains unirrigated (Ahmed et al., 1986). The growing season is characterized by low rainfall and relatively high evaporative demand. Crop grown without irrigation during the dry season thus encounters drought stress of varying degrees at different growth stages. Drought is probably the most important factor controlling crop yield worldwide. But the yield reduction is mediated through reduced leaf growth (Ephrath and Hesketh, 1991) and consequential lower photosynthetic productivity (Chen et al., 1993). The effect of drought stress on the plant growth process has been extensively reported (Hsiao, 1973; Schulze, 1986).
Drought affects nearly all the plant growth processes; however, the stress response depends upon the intensity, rate, and duration of exposure and the stage of crop growth (Brar et al., 1990). Reduction in photosynthesis in water stressed leaves may be due to stomatal closure (Hsiao, 1973). Farquhar and Sharkey (1982) gave a detailed account of the stomatal functioning in relation to stress
conditions. Recent studies indicate that a decrease in photosynthesis under deficient soil moisture may not necessarily be related to stomatal opening, rather non-stomatal control of photosynthesis might have greater influence (Farquhar et al., 1989).
This study was undertaken to investigate the photosynthetic rate and leaf gas exchange characteristics of wheat under different levels of drought stress.
Materials and Methods
An experiment was conducted under semi-controlled conditions at the Institute of Postgraduate Studies in Agriculture (IPSA), Bangladesh, between November 1994 and March 1995. Four cultivarsnamely Kanchan (V1), Sonalika (V2), Kalyansona (V3), and C306 (V4)were grown in Waggner pots (24 cm diameter) containing 12 kg sandy clay loam soil. A fertilizer dose of 600 mg N, 360 mg P, 240 mg K, and 120 mg S as urea, TSP, MP, and gypsum was applied to each pot. Ten seeds were planted per pot, and one week after emergence the seedlings were thinned to three seedlings per pot.
There were 160 pots, with 40 pots assigned to each cultivar. The plants were subjected to four levels of water regimes. These were:
1) Control. No drought stress was imposed, and pots were never allowed to dry out, (S1).
2) Vegetative. Drought stress was imposed at an early vegetative stage by withholding irrigation, (S2).
3) Anthesis. Drought stress was imposed at anthesis by withholding irrigation, (S3).
1Assistant Commissioner, Naogaon Collectorate, Naogaon, Bangladesh
2Corresponding author: Scientific Officer, Agronomy Division, Bangladesh Rice Research Institute, Gazipur-1701, Bangladesh. E-mail: enrecabd@bdonline.com or brrihq@bdonline.com