Bot. Bull. Acad. Sin. (1999) 40: 147-152
Yen et al. Sugar uptake in cultured soybean cells
Sugar uptake by photomixotrophic soybean suspension cultures1
H. Emilie Yen3, Yu-Chi Chen2, Shi-Kae Yen, and Jeng-Horng Lin
Department of Botany, National Chung-Hsing University, Taichung 402, Taiwan, Republic of China
(Received June 17, 1998; Accepted September 23, 1998)
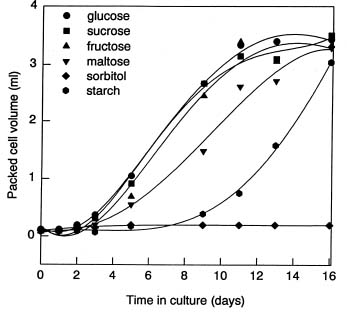
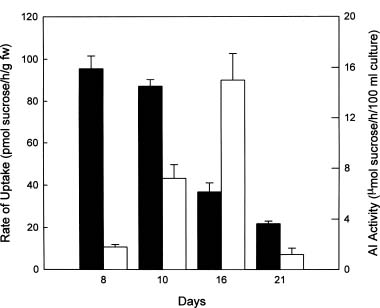
Abstract. A photomixotrophic soybean (Glycine max) culture with substantial chlorophyll content was used to study the ability of cells to grow on different carbon sources. With the exception of sorbitol, these photomixotrophic cells were able to grow on media containing 3% starch, or 2% sucrose, maltose, glucose and fructose; and the growth curves and biomass production of hexose- or disaccharide-grown cells were all fairly similar. The highest biomass production was found in the sucrose-grown cells. Levels of soluble protein and total chlorophyll and the Chl a/b ratio were highest in the starch-grown cells. Although the growth curves of sucrose-, glucose-, or fructose-grown cells were quite similar, the abilities of cells grown on one source to utilize various carbon sources were quite distinct. 14C-labelling experiments showed that the highest incorporation rates for sucrose and hexoses all occurred during the logarithmic phase with sucrose-grown cells preferentially taking up sucrose and hexose-grown cells preferentially taking up hexoses. Cells maintained on 3% starch showed an ability to take up both sucrose and hexoses with the highest uptake rate when fed with 14C-glucose. The rate of uptake of sucrose was decreased more than twofold as cells entered the stationary phase, while there was a twofold increase in the activity of extracellular acid invertase (EC 3.2.1.26). The clear correlation between low acid invertase activity and high levels of incorporated radioactivity from 14C-sucrose strongly suggested that intact sucrose is being taken up in sucrose-fed soybean cells during the vigorous growth stage, a stage similar to developing soybean pods.
Keywords: Acid invertase; Glycine max; Photomixotrophic suspension culture; Sugar uptake.
Abbreviations: AI, Acid Invertase; Chl, Chlorophyll; PA, Photoautotrophic; PM, Photomixotrophic.
Introduction
There are two known mechanisms for uptake from the apoplast of sucrose in higher plants: One is the direct uptake of exogenous sucrose (for references, see Madore and Lucas, 1989). Alternatively, sucrose is hydrolyzed extracellularly to glucose and fructose, and then retrieved by cells in the form of hexose (for references, Eschrich, 1989). In both cases, transport proteins mediate the uptake of these sugars from the extracellular space through the plasma membrane. Two kinds of carriers, namely sucrose and hexose transporters, have been identified from several plant species (Riesmeier et al., 1992; Sauer et al., 1990). The location of specific carriers in specific tissues/cells is important in the partitioning of sugars between sources and sinks (Riesmeier et al., 1994).
However, how specific cells sense sugars and what the signalling pathways are which induce specific carrier protein are still unknown. Furthermore, determination of uptake kinetics using intact leaves or leaf discs is complicated by the contribution of intercellular invertase
and the diffusive resistance of the cuticular layer. To eliminate the factor of intercellular transport, isolated protoplasts have been used extensively (Brown et al., 1997; Fieuw and Willenbrink, 1991) but only for short-term labelling studies. Photomixotrophic (PM) cells offer a system which is photosynthetically active but still requires an exogenous sugar supply and a comparatively homogeneous material for synchronous induction, growth measurements, and uptake studies. Previous studies showed that the physiological state of these cultured cells is most comparable to that of young dividing leaf cells (Rogers et al., 1987). We have maintained a soybean cell line grown on 3% starch as the major carbon source and have investigated the ability of these cells to utilize various exogenously supplied sugars as their carbon source.
Materials and Methods
Cell Culture
The cell line used in this study was originally provided by Dr. J.M. Widholm, University of Illinois. The PM callus of soybean was maintained on solid medium according to Horn et al. (1983). The suspension cells were raised by placing cells in the liquid medium containing 3% starch (Aldrich Co.) and subculturing every 3 weeks. The experiments were initiated by transferring 3 ml of 3-wk-old concentrated cells into 100 ml of liquid medium containing 2% sucrose, glucose, fructose, maltose, or
1This work in part full filled the bachelor thesis requirements for Ms. Yu-Chi Chen.
2Present Address: Department of Botany, National Taiwan University, Taipei, Taiwan.
3Corresponding author. Phone: 886-4-2840416 ext 520; Fax: 886-4-2874740; E-mail: heyen@dragon.nchu.edu.tw