Uritani Postharvest biochemistry of tropical tuberous crops
Bot. Bull. Acad. Sin. (1999) 40: 177-183
(Invited review paper)
Biochemistry on postharvest metabolism and deterioration of some tropical tuberous crops
Ikuzo Uritani
School Corp. Aichi Konan Gakuen, Konan, Aichi and Nagoya University (Prof. Emeritus), Nagoya, Aichi, Japan
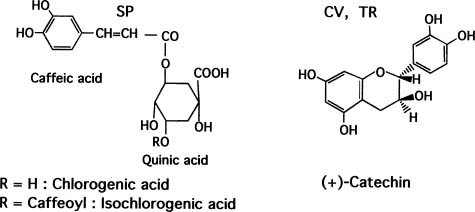
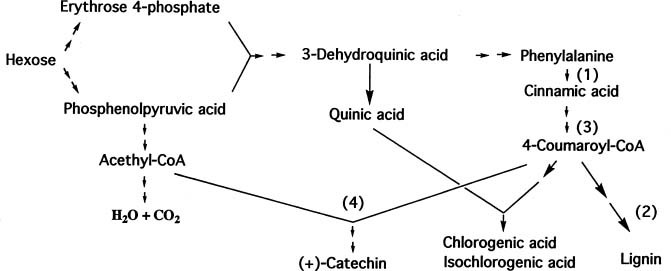
Abstract. This paper deals with a biochemical comparison of the postharvest metabolism and deterioration among three tropical tuberous crops: sweet potato, cassava, and taro, focusing on sweet potato. These often suffer from wounding and infection during harvest, transportation, and storage. In response to wounding or infection, the three crops underwent some similar metabolic alterations, such as in the production of stress compounds and changes in proteins, but they also showed some different reactions. In sweet potato and taro, wounding was often healed soon by the formation of a lignin layer succeeding production of polyphenols and changes in proteins. No such layer formed in cassava, which deteriorated at xylems soon after harvest in a phenomenon called vascular streaking, vascular discoloration, or physiological deterioration. When the three crops were attacked by microbes or insects, metabolic changes were induced more vigorously, including the induction of some enzymes, production of stress compounds such as coumarins and phytoalexins, and more enhancement of polyphenol production. However, in cassava, the stress compounds were also produced in the case of wounding.
Keywords: Colocasia esculenta; Coumarin; Ipomoea batatas; Manihot esculenta; Microbial deterioration; Physiological deterioration; Phytoalexin; Polyphenol; Storage protein; Tropical tuberous crops.
Abbreviations: SP, Sweet potato; CV, cassva; TR, taro; PD, physiological deterioration; MD, microbial deterioration; PAL, phenylalanine ammonia lyase.
Contents
Introduction 177
Materials and Classification 177
Response to Wounding 178
Response to Infection 178
Polyphenol Production 178
Production and Structures 178
Phenylalanine Ammonia Lyase and Peroxidase 179
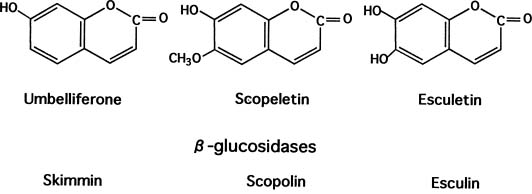
Coumarin Production 179
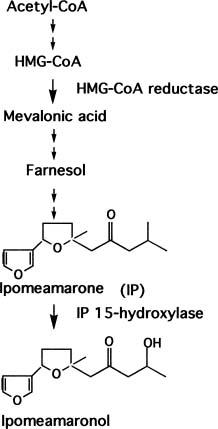
Phytoalexin Production 179
Changes in Proteins 181
Chilling Injury 181
Conclusion 181
Literature Cited 182
Introduction
There are many kinds of tuberous crops in the tropics, some of which are utilized as dietary staples. However, much scientific research on these crops remains to be performedincluding the field of postharvest biochemistry. A summary of the research which has been done, and of the questions yet to be elucidated on those crops, may be helpful in solving the world food shortage that is anticipated for the twenty-first century due to increases in the world population.
Materials and Classification
Postharvest metabolism and deterioration of sweet potato (SP, Ipomoea batatas [L.] Lam.), cassava (CV, Manihot esculenta Cranz), and taro (TR, Colocasia esculenta (L.) Schott, or C. antiquorum) after harvest are described from the biochemical point of view. The three produce tuberous organs under ground that contain large amounts of starch. All three originate in the tropical zone. However, the botanical classifications of class, family and genus, as well as chromosome and genome numbers, differ from each other (see Table 1). Hence, their natures are similar in some parts, but dissimilar in others. However, an understanding of the nature of one species of tuberous organ is useful in understanding the nature
Present address: 1-62-8, Karayama-cho, Chikusa, Nagoya, 464-0806, Japan. Tel: 81-52-781-6953; Fax: 81-52-781-6968; E-mail: iuritani@mb.infoweb.ne.jp