Kim et al. Physiological and gas emissions in rice
Bot. Bull. Acad. Sin. (1999) 40: 185-191
Physiological functions and methane and oxygen exchange in Korean rice cultivars grown under controlled soil redox
potential
J.D. Kim1, A. Jugsujinda2, A.A. Carbonell-Barrachina, R.D. DeLaune, and W.H. Patrick, Jr.
Wetland Biogeochemistry Institute, Louisiana State University, Baton Rouge, LA 70803-7511, USA
(Received April 17, 1998; Accepted January 6, 1999)
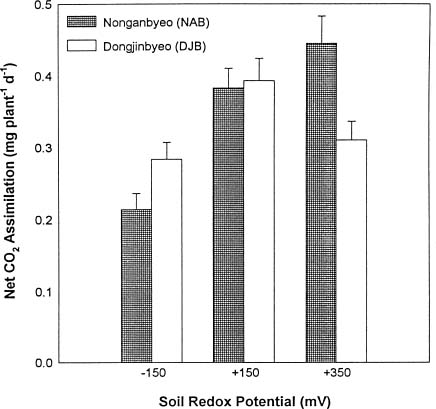
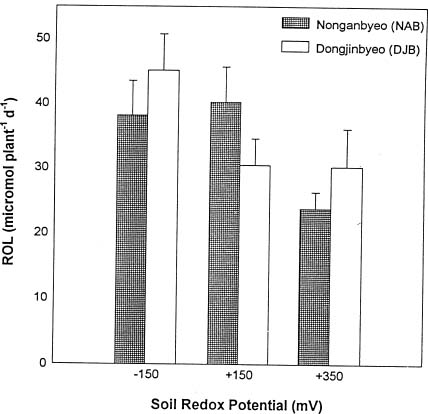
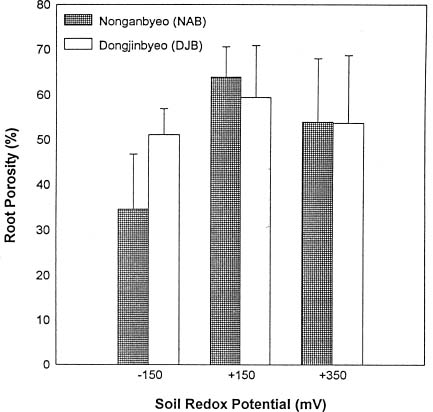
Abstract. Two Korean rice varieties (Oryza sativa L., cv. Nonganbyeo and Dongjinbyeo) were grown under laboratory conditions in Crowley silt loam (fine, montmorillonitic, thermic fluvaquent) soil suspension maintained at different redox potential (Eh) levels (-150, +150 and +350 mV). Plant growth, aerenchyma formation, radial oxygen loss, net CO2 assimilation, CH4 production (soil) and emission were measured. In general, plant height, root length, dry weight and carbon fixation for Nonganbyeo decreased under strongly reduced conditions (-150 mV). For Dongjinbyeo only a decrease in CO2 fixation was observed at -150 mV as compared to moderately reduced conditions (+150 mV and +350 mV). Under strongly reduced conditions (-150 mV) root porosity was less than rice grown at the two higher redox levels. Since greatest soil methane production occurred under strongly reduced conditions (-150 mV) no emission from the two rice cultivars were measured under moderately reduced soil conditions. However, under the strongly reduced soil conditions where methane was formed, there were apparent differences in methane emission between the two varieties studied. Methane production in pot in which DJB was grown was approximately 50 times greater than that of NAB but net emission was only greater by the factor of 3. Net carbon assimilation varied depending on soil Eh. Reduced soil conditions (Eh -150 mV) decreased net photosynthesis for both varieties. Radial oxygen loss on the other hand was greatest at Eh -150 mV, and this can be attributed to more oxygen demand under more reduced soil conditions. Results demonstrate that the degree of soil reduction influences both rice growth and methane emission. Management practices should consider growing the two rice cultivars under moderately reduced soil conditions where best carbon fixation was measured. Growth under these conditions would also reduce methane emission to atmosphere.
Keywords: Aerenchyma formation; Carbon dioxide fixation; Korea; Methane emission; Oryza sativa; Radial oxygen loss.
Abbreviations: DJB, Dongjinbyeo; Eh, redox potential; NAB, Nonganbyeo; NCA, net CO2 assimilation, ROL, radial oxygen loss.
Introduction
Rice is vital to more than half of the world's population (Vlek and Byrnes, 1986). It is the most important food grain in the diet of hundred of millions of Asians, Africans, and Latin Americans living in the tropics and subtropics. Approximately 75~80% of cultivated rice is grown under rainfed or irrigated wetland conditions or under submerged paddy field conditions. This is done to increase grain yield (Lindau et al., 1993) or because biophysical conditions do not allow the growth of crops upland.
Soil redox potential can affect rice growth. Subsequent to flooding, plant roots encounter hypoxic and anoxic conditions as a result of the depletion of soil oxygen by microbial and plant root respiration (Gambrell and Patrick,
1978). Flooded soils can experience redox conditions ranging from well-oxidized to strongly reduced. In an oxidized (aerobic) soil the redox potential (Eh) ranges from about +600 to +350 mV, whereas in most reduced (anaerobic) soils the Eh varies from about -300 to +350 mV (DeLaune et al., 1990; Masscheleyn et al., 1993).
Methane gas is one of the greenhouse gases that is reported to exert significant effects on the global heat balance, thus causing a possible elevation of global surface temperature (Bouwman, 1991). Rice paddies can serve as CH4 sources and sinks because soil conditions allow methane-generating and methane-oxidizing bacteria to exist in close approximation (Oremland and Culbertson, 1992). CH4 emissions from paddy fields are highly variable and determined by complex interactions among the chemical, physiological, and biological properties of the local environment (Bartlett et al., 1985). Rice plants develop aerenchyma, which provides the roots with O2 (Kludze et al., 1994). Thus, much higher CH4 emissions were found in vegetated rice fields than in those that were not vegetated (Cicerone and Shetter, 1993).
1Present Address: National Honam Agricultural Experiment Station, R.D.A., P.O. Box 5, Iksan, 570-080, Republic of Korea.
2Corresponding author. Tel: (504) 388-6430; Fax: (504) 388-6423; E-mail: ajugsujind@aol.com