Hsu et al. C and N compounds in flooded wax-apple trees
Bot. Bull. Acad. Sin. (1999) 40: 193-198
The fluctuation of carbohydrates and nitrogen compounds in flooded wax-apple trees
Yu-Mei Hsu1, Menq-Jiau Tseng2 and Chin-Ho Lin1,3
1Department of Botany and 2Department of Horticulture, National Chung Hsing University, Taichung, Taiwan 40227, Republic of China
(Received May 21, 1998; Accepted October 28, 1998)
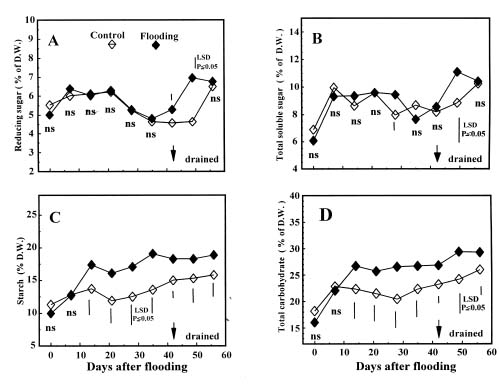
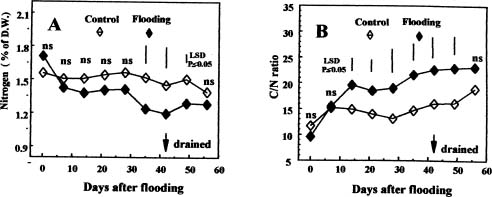
Abstract. The present study associates the response to flooding of potted wax-apple trees, Syzygium samarangense Merr. et Perry, with changes in carbon and nitrogen metabolism. In wax-apple leaves, the starch content conspicuously increased after 14 days of flooding, and the total nitrogen content decreased after 35 days of flooding. In roots, concentrations of total soluble sugars significantly increased after 14 days of flooding. The accumulation of starch in flooded wax-apple leaves and increased soluble sugar content in roots apparently were the result of the reduction of growth and metabolic activities in roots after flooding, which reduced the sink demand of carbohydrates. Soluble protein concentration significantly decreased after 7 days of flooding, but remained similar among 14-, 28- and 42-day flooding treatments. Free amino acid content of flooded plants was significantly higher than that of the control at all sampling dates. The activity of glutamine synthetase in leaves significantly decreased after 7 days of flooding, but was higher than the control`s at 14, 28 and 42 days of flooding. In wax-apple roots, content of soluble protein, free amino acids, ammonia and the activity of nitrate reductase and glutamine synthetase all decreased significantly after flooding. Apparently, nitrogen metabolism was restricted in the roots during flooded conditions.
Keywords: Carbon/nitrogen ratio; Free amino acid; Glutamine synthetase; Nitrate reductase; Nitrogen; Soluble protein; Soluble sugar; Starch; Syzygium samarangense.
Introduction
In general, flooding has an adverse effect on fruit trees. In Taiwan, constant flooding of wax-apple tree (Syzygium samarangense Merr. et Perry) for 45 days during the summer not only advances the harvest period to the following December, but also increases harvest intervals and productivity (Wang, 1983). Long-term flooding results in soil anoxia, which restricts root growth and causes a metabolic imbalance between shoots and roots. Zvareva and Bartkov (1976) demonstrated that flooding decreased carbohydrate translocation from the leaves to the roots of soybean. Similar effects have also been found in Saxifraga tormentosa (Quereshi and Spanner, 1973), purple flower alfalfa (Medicago sativa L.) and Lotus corniculatus L. (Barta, 1987). The reduction of photosynthate translocation to roots under flooding stress might have been due to the reduction of carbohydrate utilization in roots (Wample and Davis, 1983) or to depression of the photosynthate transport system (Saglio, 1985; Topa and Cheeseman, 1992a). Therefore, under flooding conditions, although the photosynthetic rate in leaves declines, starch accumulates (Barta, 1987; Topa and Cheeseman, 1992b; Vu and Yelenosky, 1991; Wample and Davis, 1983).
Restriction of the absorption of mineral nutrients in roots under flooding usually results in lower nitrogen content in the tissues of flood-intolerant plants (Kozlowski and Pallardy, 1984). Oxygen deficiency restricts protein synthesis in roots (Lin and Key, 1967; Sachs et al., 1980) and accelerates anoxic metabolism (Crawford and Baines, 1977; Jackson et al., 1982). Therefore, under flooding stress, the composition and quantity of proteins and amino acids, and the activities of key enzymes involved in nitrate reduction and ammonia assimilation, will all be affected (Buwalda et al., 1988; Garcia-Novo and Crawford, 1973; Reggiani et al., 1988).
Wax-apple tree, a flood-tolerant species, is expected to have a different carbon and nitrogen metabolism response to flooding than flood-intolerant plants. Thus, the present study analyzed the variation of carbon and nitrogen contents in the leaves and roots with and without flooding. It also evaluated the impact of flooding on the activities of nitrate reductase and glutamine synthetase.
Materials and Methods
Plant Preparation and Flooding Treatments
Experiments were performed under open field conditions without fertilization on three-year-old wax-apple plants (Syzygium samarangense Merr. et Perry), grown in 16-L non-woven fabric bags with bark compost as a growth medium. Eighteen bagged plants were randomly divided
3Corresponding author. Tel: 886-4-2862511; Fax: 886-4-2862511; E-mail: chlin@mail.nchu.edu.tw