Wang et al. A desiccation-related protein in lily pollen
Bot. Bull. Acad. Sin. (1999) 40: 199-205
A stress-inducible protein associated with desiccation in lily pollen
C.-S. Wang1, S.-M. Lin, and S.-L. Wei
Graduate Institute of Agricultural Biotechnology, National Chung Hsing University, Taichung, Taiwan, Republic of China
(Received June 11, 1998; Accepted November 24, 1998)
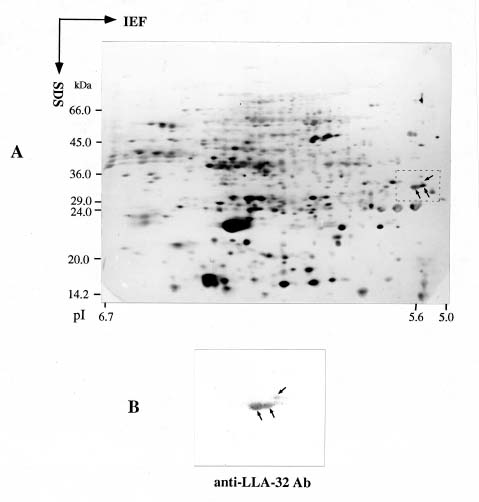
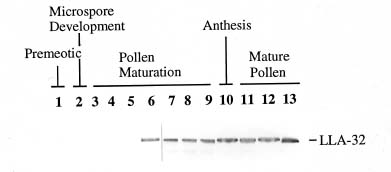
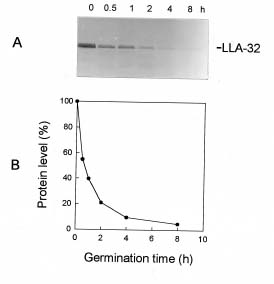
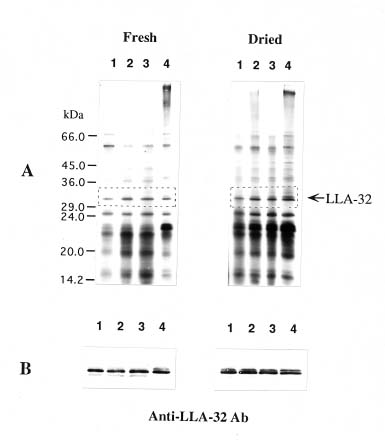
Abstract. This work characterizes a stress-inducible protein (LLA) associated with desiccation in lily (Lilium longiflorum Thunb. cv Snow Queen) pollen. Immunoblots of two-dimensional polyacrylamide gels of pollen protein indicated that the LLA-32 protein with a molecular mass of 32 kDa was heterogeneous and had an isoelectric point of 5.6. The protein was developmentally regulated at the later stage of pollen maturation during anther development. The accumulation of LLA-32 proteins remained steady in the mature pollen, but the protein disappeared during pollen germination. Subcellular fractionation of pollen proteins revealed that the protein was detected only in the cytoplasmic fraction, indicative of a gametophytic origin. Premature drying of developing pollen demonstrated that the LLA-32 doublet was associated with desiccation. The accumulation of the proteins can be experimentally manipulated by methyl jasmonate and salicylic acid as well as by mannitol and methyl viologen. Differential accumulation of the doublet polypeptides occurred with response to various stimuli. The upper band of LLA-32 doublet polypeptides markedly accumulated upon dehydration whereas the lower band of LLA-32 doublet significantly increased its level of accumulation when other stresses were applied.
Keywords: Desiccation; Lilium longiflorum; Pollen protein; Stress-inducible.
Introduction
Anthers are floral structures specialized for pollen production in higher plants. In young anthers, the sporogenous cells undergo meiosis and give rise to the formation of microspores at the tetrad stage. These microspores develop into microgametophytes after haploid mitosis and subsequently differentiate into mature pollen (Johri, 1984). In the final stage of maturation, pollen exhibits various degrees of desiccation and this natural drying process promotes pollen germinability to a great extent.
As an adaptive organ that facilitates the colonization of terrestrial habitats, pollen develops a number of unique features (Blackmore and Knox, 1990). First, the pollen grain is surrounded by a protective wall consisting of two layers, exine and intine. Second, sporopollenin in the exine is an insoluble polymer that is extremely resistant to drought and chemicals, including strong acids. Finally, the presence of soluble sugars such as sucrose in pollen may play an essential role in the acquired tolerance to desiccation (Hoekstra and van Roekel, 1988). Anhydrobiotic organisms are known to contain trehalose, a nonreducing disaccharide of glucose that is related to acquired toler
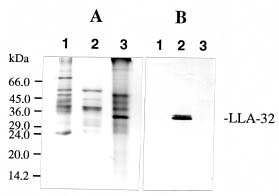
ance to severe dehydration (Crowe et al., 1984). In addition to sugars, proteins are considered to be another component of desiccation tolerance. Various types of drought-inducible proteins have been identified (Skriver and Mundy, 1990; Shinozaki and Yamaguchi-Shinozaki, 1997). Among these, dehydrins are known to accumulate immediately before desiccation during seed development (Close et al., 1989; Dure, 1993). Dehydrins are also reported in mature pollen grains (Michel et al., 1994; Wang and Cutler, 1995). It has been suggested that these proteins play a role in protecting plant structures during water loss. Most genes that respond to drought are also abscisic acid (ABA)-responsive (Ingram and Bartels, 1996; Bray, 1997). We recently described two pollen-specific proteins that were immunologically unrelated to dehydrins in lily seeds (Wang et al., 1996, 1998). The two pollen proteins that accumulated shortly before anthesis were ABA and polyethylene glycol (PEG)-inducible (Wang et al., 1996). Herein, we continue our examination by using immunological and biochemical methods to characterize one of the two proteins during development and stress. The LLA-32 protein is a heterogeneous protein located in the cytoplasmic fraction of pollen grains. Premature drying of developing pollen demonstrates that the doublet polypeptides are associated with desiccation. The LLA-32 doublet polypeptides differentially increase their levels of accumulation upon dehydration and other stresses.
1Corresponding author. Tel: 886-4-287-4754 ext. 105, 106; Fax: 886-4-286-1905; E-mail: cswang2@dragon.nchu.edu.tw