Wang et al. Osmotic stress induced metabolic changes
Bot. Bull. Acad. Sin. (1999) 40: 219-225
Effect of sorbitol induced osmotic stress on the changes of carbohydrate and free amino acid pools in sweet potato cell suspension cultures
Heng-Long Wang1,2, Ping-Du Lee2, Li-Fei Liu3 and Jong-Ching Su1,2,4
1Institute of Biological Chemistry, Academia Sinica, P. O. Box 23-106, Taipei, Taiwan 107, Republic of China
2Biochemistry Laboratory, Department of Agricultural Chemistry, National Taiwan University, 1 Roosevelt Road Section 4, Taipei, Taiwan 107, Republic of China
3Department of Agronomy, National Taiwan University, 1 Roosevelt Road Section 4, Taipei, Taiwan 107, Republic of China
(Received November 23, 1998; Accepted January 5, 1999)
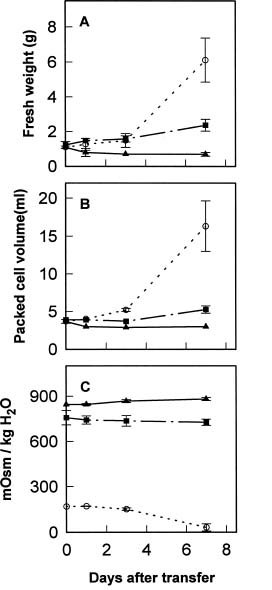
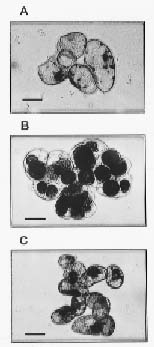
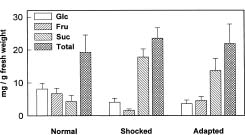
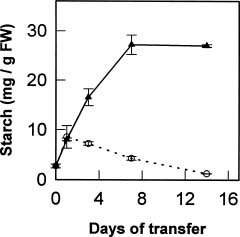
Abstract. The effects of osmotic stress induced by 0.6 M sorbitol on the cell growth and on the quantitative and qualitative changes in carbohydrates and free amino acids in suspended cells of sweet potato (Ipomoea batatas) were analyzed. Cells transferred into medium without (normal treatment) or with (stress-shocked treatment) 0.6 M sorbitol added, and cells consecutively subcultured under high stress conditions (stress-adapted treatment) were compared. Stress-shocked cells showed cell growth retardation and the induction of plasmolysis. Stress-adapted cells had a shorter lag phase in growth than the stress-shocked, and showed a normal morphology, albeit the size appeared slightly smaller than normal cells. Under the stress-shocked condition, the size of the amino acid pool (µmole/g fresh weight) increased fourfold relative to the control and stress-adapted cells. The levels of alanine and glutamic acid and its derivatives were especially high, indicating that the changes in the intensity of glycolysis have influenced the amino acid pool. Although the proline level showed a fivefold increase when stress-shocked, proline made up only about 1.5% of total amino acids, and thus did not seem to play an osmotic regulatory function. Among the carbohydrates, sucrose content was high in both stress-shocked and stress-adapted cells. Starch accumulated heavily in stress-shocked cells, but not in normal or stress-adapted cells, although the latter maintained a higher background level of starch. It is tempting to speculate that sucrose serves as a compatible solute, and starch synthesis from sucrose plays a pivotal role in moderating the hyperosmotic condition. The accumulated starch contained less amylose than the ordinary tuberous root starch, indicating that the pathway of starch synthesis was somewhat altered in the stress-shocked cells.
Keywords: Compatible solute; Free amino acid; Ipomoea batatas; Osmotic stress; Starch; Sucrose; Suspended cells.
Abbreviations: 2,4-D, 2,4-dichlorophenoxyacetic acid; GABA, g-aminobutyric acid; MS, Murashige-Skoog.
Introduction
Plants resort to many adaptive strategies in response to abiotic environmental stresses such as high salt, dehydration, cold, heat, and excessive osmotic pressure. These adaptive mechanisms include changes in morphological and developmental patterns as well as physiological and biochemical processes (McCue and Hanson, 1990). Among them, the accumulation of compatible solutes according to the metabolic responses has drawn much attention. Some stress-responsive genes encoding proteins for compatible solute synthesis have been cloned and expressed in transgenic plants (Tarczynski et al., 1993; Bartels and Nelson, 1994; Kavi Kishor et al., 1995). The compatible solutes may be classified into two categories:
one is nitrogen-containing compounds such as proline and other amino acids, quaternary ammonium compounds and polyamines, and the other is hydroxy compounds, such as sucrose, polyhydric alcohols, and oligosaccharides (McCue and Hanson, 1990). The species of accumulated solutes varies with the variation in adverse conditions and plant species, or even plant varieties.
In general, a plant cell suspension culture is considered a relatively homogeneous population of cells. Much research has used cultured cells as a model system to study the cellular responses under various abiotic stress, even to distinguish the difference between the short-term response and long-term adaptation involving physiological and biochemical changes (Fallon and Phillips, 1989; Leone et al., 1994).
Sorbitol is an alditol found in higher plants. It is the major photoassimilate in most species of Rosaceae (Moing et al., 1992) and the main low molecular weight saccharide
4Corresponding author. E-mail: jcs@gcms.ntu.edu.tw