Bot. Bull. Acad. Sin. (1999) 40: 251-257
Thseng et al. Glycine formosana in Taiwan
Glycine formosana Hosokawa in Taiwan: pod morphology, allozyme, and DNA polymorphism
F.S. Thseng1, 3, S.J. Tsai1, J. Abe2, and S.T. Wu1
1Department of Agronomy, National Chung Hsing University, Taichung 402, Taiwan
2Laboratory of Plant Genetic Resources, Hokkaido University, Japan
(Received April 22, 1998; Accepted January 15, 1999)
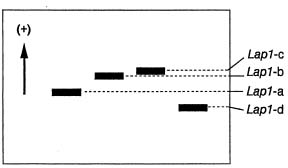
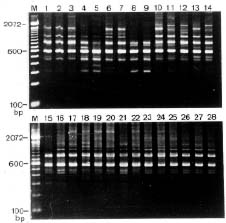
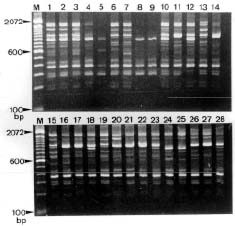
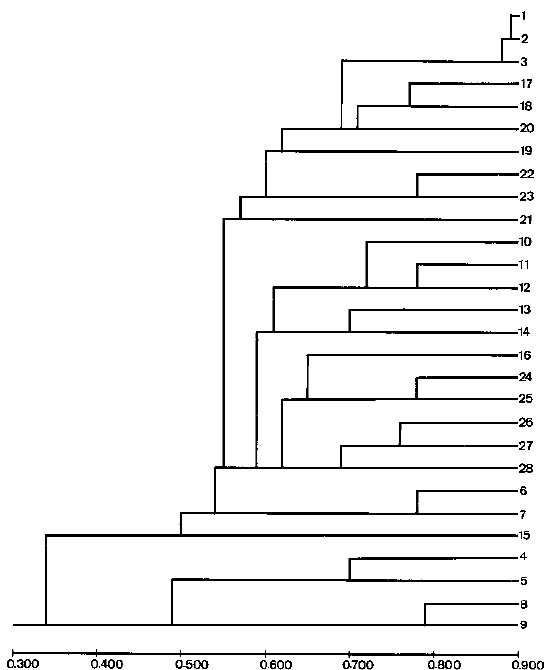
Abstract. Glycine formosana Hosokawa is distributed over the grassland along the riverside and roadside at Dahshi, Guanshi, and Herngshan in Taoyuan county, Taiwan. It is a twining annual herb. It was long considered to be of the same species as G. soja, distributed widely in East Asia. Because the plant showed a continuous variation with G. soja in appearance, Tateishi and Ohashi (1992) considered it a geographic subspecies and renamed it G. max subsp. formosana (Hosokawa) Tateishi et Ohashi. This study attempts to determine the differences between G. formosana and the G. soja collected in China, Korea, and Japan. The materials were planted at National Chung-Hsing University. The pods, allozymes, and DNA polymorphisms were investigated. Glycine formosana has small seeds which are significantly different from the G. soja in China, Korea, and Japan. Lap1-d one of the 16 loci of 9 allozymes exists only in G. formosana. Twenty-five random sequence 10-mer primers were employed in RAPD analysis for all samples. Twenty-one produced bands, and 14 of those showed polymorphisms. One hundred and thirty-two bands were produced, and 84 bands (64%) showed polymorphisms. Based on the appearances of markers, the genetic similarity coefficients were calculated. Among different samples of G. formosana, few genetic variations were observed (0.885~0.887). However, G. formosana showed marked differences from G. soja in China and Korea and nested within the Japan accessions.
Keywords: Allozyme; DNA polymorphism; Glycine formosana; G. soja; Pod; Seed.
Abbreviation: RAPD, random amplified polymorphic DNA.
Introduction
Glycine consists of many species in two subgenera, Soja and Glycine. Subgenus Soja includes G. max, a cultivated form, and G. soja, a wild relative. Subgenus Glycine has at least 17 species including G. albicans, G. arenaria, G. argyrea, G. canescens, G. clandestina, G. curvata, G. cytoloba, G. dolichocarpa, G. falcata, G. hirticaulis, G. lactovirens, G. latifolia, G. latrobeana, G. microphylla, G. pindanica, G. tabacina, and G. tomentella (Tindale, 1984, 1986a,b; Tindale and Craven, 1988, 1993; Tateishi and Ohashi, 1992).
Glycine soja is generally distributed in China, Japan, Korea, and Taiwan (Hymowitz and Newell, 1981). In 1924, Shimada collected a wild relative in Hsinchu and Dahshi, Taiwan, and recognized it as G. ussuriensis Regal et Maack (cf: Tang and Lin, 1962). Soon after, another wild relative was collected in Hsinchu, Chutong, and Herngshan. In 1932, Hosokawa identified and classified the latter as G. formosana Hosokawa. Not until 1962 did Tang, Lin, and Hermann identify the two wild relatives as the same species as G. soja. Huang and Ohashi (1977) replaced the name G. ussuriensis with G. soja Sieb. et Zucc.
In the mid-1980s, Ohashi et al. (1984) again replaced it with G. max subsp. soja (Sieb. et Zucc.) Ohashi. Recently, considering the characteristics of its leaf, flower, pod, and seed, Tateishi and Ohashi (1992) named this wild relative found in Taiwan as G. max subsp. formosana (Hosokawa) Tateishi et Ohashi. However, all of the above were based on the result of plant morphology investigations. In contrast, this experiment is based on DNA and allozyme investigations.
Materials and Methods
Plant Materials
Three accessions (accession numbers 1~3) of G. formosana were collected from three counties in Taiwan, including Dahshi, Guanshi, and Hernshan. Each accession collected ten plants as a population from each county. Two accessions of G. soja were collected from Korea (accession numbers 4~5), four from China (accession numbers 6~9), and nineteen from Japan (accession numbers 10~28). Twenty seeds were randomly selected from each accession. The seeds served as base materials in this experiment. First, their coatings were pierced, and then the seeds were planted on plastic plates. When the seedlings had grown to about three centimeters, they were moved to pots; two in each pot and ten pots for each
3Corresponding author. Fax: (04) 286-2171.