Bot. Bull. Acad. Sin. (1999) 40: 259-265
Yang and Wang Protease and amylase production of Streptomyces
Protease and amylase production of Streptomyces rimosus in submerged and solid state cultivations
Shang-Shyng Yang1,3 and Jan-Yi Wang2
1Department of Agricultural Chemistry, National Taiwan University, Taipei 106, Taiwan
(Received May 21, 1998; Accepted February 12, 1999)
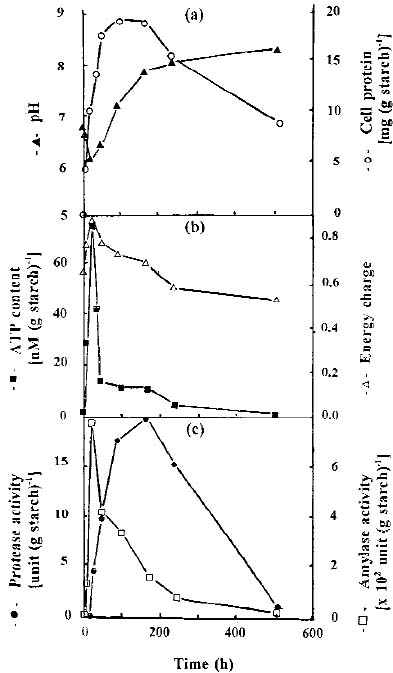
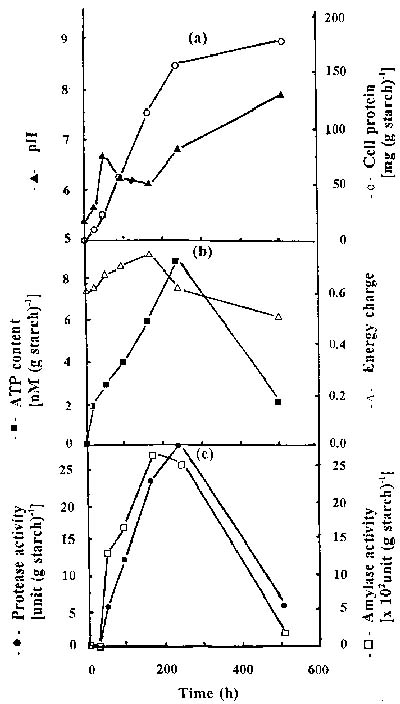
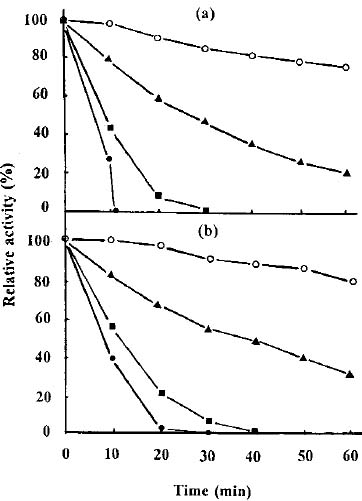
Abstract. The protease activity of Streptomyces rimosus TM-55 was first detected after 12 h of growth in submerged cultivation, and this activity peaked after 166 h of incubation. In solid state cultivation, protease was first secreted at 24 h, with the secretion peaking at 232 h. Amylase activity could be detected after 6 h in submerged cultivation, and it afterwards peaked after 48 h of incubation. In solid state cultivation, it began to be secreted at 24 h, and this secretion peaked after 180 h. Each gram of starch yielded 17.4 and 691.3 units of protease and amylase in submerged cultivation, respectively; whereas the values were 26.7 and 2,642.7 units in solid state cultivation. a-Amylase was the major amylase in both cultivation methods, and glucoamylase and debranching activity were minor components. Protease and amylase produced with both cultivation methods had a similar optimal pH, between 6.0 and 7.0, and optimal temperature, between 35 and 45°C. The enzyme activities produced in solid state cultivation were more stable with pH and temperature changes than those produced in submerged cultivation.
Keywords: Amylase; Protease; Streptomyces; Submerged and solid state cultivations.
Introduction
In Taiwan and other Asian countries, the Koji process has been used to produce various enzymes by growing molds on cereals or their brans. Although protease and amylase are mainly fungal and eubacterial products, the possibility of using streptomycetes for enzyme production has recently been investigated. Streptomyces species that produce proteases include S. clavuligerus, S. griseus, S. moderatus, S. rimosus, S. thermoviolaceus, and S. thermovulgaris (Pokorny et al., 1979; Renko et al., 1981, 1989; Chandrasekaran and Dhar, 1987; Bascaran et al., 1990; James et al., 1991; Muro et al., 1991; Yeoman and Edwards, 1994). Other hydrolases of Streptomyces species studied include aminopeptidase by S. fradiae, S. griseus, S. lividans, S. peptidofaciens, and S. rimosus (Vitale et al., 1986; Aphale and Strohl, 1993), chitinase by S. viridificans (Gupta et al., 1995), a-amylase by S. aureofaciens and S. rimosus (Vukeli et al., 1992; Cheng and Yang, 1995; Yang and Cheng, 1996), and b-glucosidase by Streptomyces sp. (Ozaki and Yamada, 1991).
Streptomyces species are heterotrophic feeders, and they can utilize both simple and complex molecules as nutrients. About three-fourths of the Streptomyces species may produce antibiotics. In addition to antibiotics, Streptomyces species liberate extracellular enzymes (Gupta et al., 1995). This work investigates the effect of sub
ú
c
merged and solid state cultivation techniques on growth, ATP content, and on the protease and amylase activities of Streptomyces rimosus.
Materials and Methods
Test Organism
Streptomyces rimosus TM-55 (CCRC 940061) was obtained from Dr. Thomas H. H. Ku, Cyanamid Taiwan Corporation.
Sweet Potato Residue and Peanut Meal Residue
Sweet potato residue and peanut meal residue were purchased from a local market in Taiwan and passed through a sieve (4 to 16 mesh) to remove dust and large aggregates. Sweet potato residue contained (w w-1) 14.0 to 16.1% moisture, 2.3 to 3.1% crude protein, 2.7 to 3.6% ash, 16.1 to 18.0% crude fibre, and 65.4 to 70.0% carbohydrate (mainly starch) (Yang, 1988). Peanut meal residue comprised (w w-1) 9.2 to 10.0% moisture, 49.4 to 51.4% crude protein, 5.4 to 6.1% ash, 6.7 to 7.5% crude fibre, 1.3 to 1.6% crude fat, and 25.2 to 26.2% nitrogen free extract.
Culture Media and Growth Conditions
Streptomyces rimosus was cultivated at 28°C on a slant containing (g l-1) soluble starch 10; yeast extract, 1; beef extract, 1; tryptone, 2; FeSO47H2O, 0.1; and agar, 20; at pH 7.2. Spores were harvested with a Tween 80 solution (5 ml, 0.05% v v-1), which was then adjusted with sterile water to give 107 to 108 spores per ml.
2Present address: National Laboratories of Foods and Drugs, Department of Health, Executive Yuan, Taipei 110, Taiwan.
3Correspondence author. Tel: 886-2-23621519; Fax: 886-2-23679827; E-mail: ssyang@ccms.ntu.edu.tw