Bot. Bull. Acad. Sin. (1999) 40: 267-273
Chen and Kuc' Ca2+-dependent excretion of salicylic acid
Ca2+-dependent excretion of salicylic acid in tobacco cell
suspension culture
Hsien-Jung Chen1 and Joseph Kuc' 2,3
1Postdoctorate, Institute of Botany, Academia Sinica, Nankang, Taipei 11529, Taiwan
2Professor Emeritus, Department of Plant Pathology, University of Kentucky, Lexington, Kentucky 40546, USA
(Received June 19, 1998; Accepted February 12, 1999)
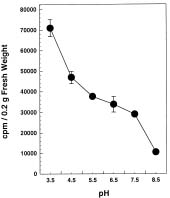
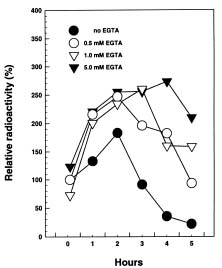
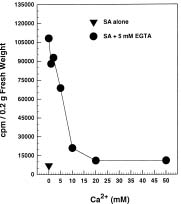
Abstract. The radioactive 14C-Salicylic acid (SA) was used to monitor the metabolism of exogenous SA in tobacco cell suspension culture. Tobacco cells took up ca. 39.46% of the applied SA 5 min after the addition of a final 200 µM SA mixture containing [14C]SA to the cell suspension culture. Most of the SA absorbed by the cells was gradually lost to the culture medium, and 2.62% and 91.55% of the radioactivity were found in the cells and the culture filtrate, respectively, 5 h after treatment. Excretion of SA to the culture medium required external Ca2+ and was inhibited by EGTA pretreatment. The inhibition was reversible through the supplement of Ca2+ to the cell suspension culture. Based on the data, we conclude that SA excretion from the 200 µM-treated cells requires external Ca2+ and likely involves a putative Ca2+-dependent pathway in tobacco suspension culture.
Keywords: Ca2+; EGTA; SA loss; Salicylic acid; Tobacco cell suspension culture.
Introduction
SA, 2-hydroxybenzoic acid, is endogenous in many plants including tobacco, cucumber, rice, wheat, cotton, tomato, and Arabidopsis (Raskin et al., 1990). SA has been considered a natural growth regulator with numerous functions in plants (Raskin, 1992). It promotes bud formation and growth in tobacco callus culture (Lee and Skoog, 1965); induces flowering in Lemna gibba G3 and Lemna paucicostata 151 (Cleland and Ajami, 1974; Tanaka et al., 1979; Watanable and Takimoto, 1979); inhibits ethylene biosynthesis in pear cell suspension culture (Leslie and Romani, 1986); inhibits jasmonic acid biosynthesis and prevents wound inducible proteinase inhibitor gene expression in tomato (Pena-Cortes et al., 1993); regulates heat production in the inflorescence of an Arum lily (Raskin et al., 1987; Raskin et al., 1989); reverses ABA-induced stomatal closure (Rai et al., 1986); induces local resistance to TMV in Nicotiana tabacum cv. xanthi-nc, Samsun NN and White Burley (White, 1979); and functions as a factor for systemic acquired resistance (SAR) in tobacco (Malamy et al., 1990; Gaffney et al., 1993; Vernooij et al., 1994a; Vernooij et al., 1994b), cucumber (Metraux et al., 1990), and Arabidopsis (Delaney et al., 1994).
The concentration of endogenous SA varied significantly from tissue to tissue, and also from species to species. In thermogenic plants such as Dioon hildebrandtii, the amount of SA could be as high as 100 µg g-1 fresh weight in male cones. However, it could be
as low as less than 0.01 µg g-1 fresh weight in leaves of Nicotiana tabacum and Zea mays (Raskin et al., 1990). In plant-microbe interactions, the endogenous SA concentration could increase 10 to 20 fold in TMV-infected leaves (Malamy et al., 1992). SA can be found in nature in both free and conjugate forms during plant development (Cooper-Driver et al., 1972) and plant-microbe interactions (Malamy et al., 1992).
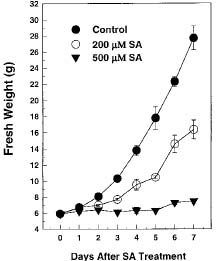
SA has been applied to tobacco cell suspension culture to study its effects on gene expression and cell physiology. Kapulnik et al. (1992) reported that 20 µM SA could induce cyanide-resistant respiration of tobacco cells in suspension culture. An SA-inducible gene encoding a 48 kDa MAP kinase protein has been isolated from tobacco. SA induction of the MAP kinase mRNA is dose-dependent with concentrations more than 50 µM, and the maximal induction could be found at the concentration of 500 µM in tobacco cell suspension (Zhang and Klessig, 1997). Kawano et al. (1998) reported that 500 µM SA could induce extracellular superoxide generation followed by an increase in cytosolic calcium ion in tobacco cell suspension culture. The acidic b-1,3-glucanase mRNA was induced by 200 µM and 20 µM SA in tobacco cell suspension culture (Chen, 1995). Therefore, the SA dose within 500 µM is appropriate and can be used to study its effects on particular gene induction and cell physiology in tobacco cell suspension culture.
In order to achieve the SA functions described above, SA has to enter the cells preceding the trigger of a particular gene expression. Ben-Tal and Cleland (1982) reported that Lemna gibba G3 plants took up nearly 90% of the applied 10 µM SA containing [14C]SA within 30 min. Rapid uptake of SA was also reported in tobacco suspen
3Corresponding author. Current address: 700 Front St., #1202, San Diego, CA 92101, U.S.A.