Bot. Bull. Acad. Sin. (1999) 40: 275-281
Pan et al. Over-expression of rice CuZnSOD in E. coli
Over-expression and characterization of copper/
zinc-superoxide dismutase from rice in Escherichia coli
Shu-Mei Pan1, Guan-Bor Hwang, and Hung-Chi Liu
Department of Botany, National Taiwan University, Taipei, Taiwan, Republic of China
(Received August 10, 1998; Accepted March 3, 1999)
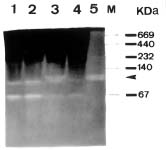
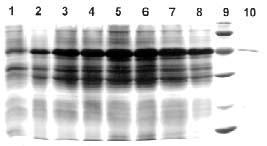
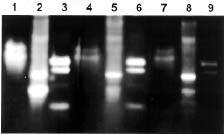
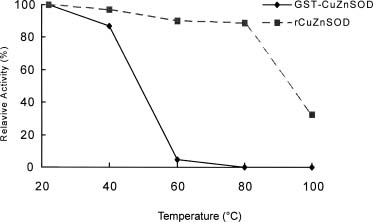
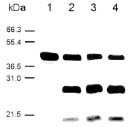
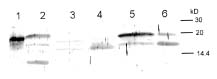
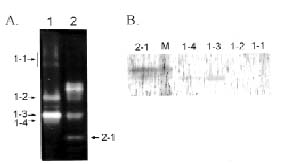
Abstract. The over-expression and characterization of cytosolic Cu/Zn-superoxide dismutase (CuZnSOD) from rice in Escherichia coli have been achieved. The full cDNA sequence coding for the rice cytosolic CuZnSOD was made by PCR and inserted into a pGEX-2T expression vector. The recombinant DNA was transformed to E. coli XL1 blue. Transformed E. coli expressed GST-CuZnSOD at levels greater than 20% of soluble protein under optimized conditions, and 10-30 mg of fusion protein can be purified from 1L bacterial culture by affinity gel. The purified fusion protein was cleaved to remove GST and produced recombinant CuZnSOD (rCuZnSOD). The MWs of a subunit of the fusion protein and rCuZnSOD were 43 kDa and 18 kDa, respectively, as predicted. The SOD activity was retained as the dimer for both forms. The fusion protein and rCuZnSOD were characterized for thermostability and the effects of pH and SDS on its activity by 10% nondenaturing gel. They showed resistance to the inhibition of hydrogen peroxide, in contrast to the native form of the plant CuZnSOD. Antiserum prepared from the GST-CuZnSOD fusion protein showed cross-reactivity to the subunits of rice cytosolic and plastidic CuZnSOD, and to those of other plants.
Keywords: CuZnSOD; Escherichia coli; Oryza sativa; Superoxide dismutase.
Abbreviations: CuZnSOD, copper/zinc superoxide dismutase; GST, glutathione transferase; PVDF, polyvinylidene difluoride; rCuZnSOD, recombinant CuZnSOD; SOD, superoxide dismutase.
Introduction
Reactive oxygen speciesincluding superoxide, hydrogen peroxide, and hydroxyl radicalsare generated by a number of pathways (Fridovich, 1978). Accumulation of these active oxygen species in aerobic organisms may cause peroxidation of membrane lipids, DNA strand breakage, and inactivation of enzymes (Imlay and Linn, 1988). Defense systems derived from enzymatic and nonenzymatic antioxidants in living organisms can minimize the deleterious effects of active oxygen free radicals (Pell and Steffen, 1991). Superoxide dismutases (SOD), catalase, and ascorbate peroxidase are among the important antioxidant enzymes in plants. SODs are a group of metal-containing enzymes that catalyze the dismutation of superoxide radical to molecular oxygen and hydrogen peroxide (Beyer et al., 1991). These enzymes may be classified into three types according to their metal cofactor requirements: copper/zinc (CuZnSOD), manganese (MnSOD), and iron (FeSOD) forms. Isoforms of CuZnSOD, the most abundant SOD in higher plants, are located both in cytosol and in chloroplasts (Kanematsu and Asada, 1989). SOD activity in plants increases differentially in re
sponse to various environmental stresses (Bowler et al., 1989; Tsang et al., 1991). SOD activity often correlates with the severity of stress (Bowler et al., 1992). It has been reported that transgenic plants having high SOD activity enhance the tolerance towards oxidative stress (Bowler et al., 1991; Perl et al., 1993; Sen Gupta et al., 1993) and other stresses (Mckersie et al., 1993; Van Camp et al., 1994), although not all highly expressing plants are oxidative stress tolerant (Tepperman and Dunsmuir, 1990). Other enzyme activities may be needed at the same time (Foyer et al., 1994).
The cDNA encoding cytosolic CuZnSODs from several plants and rice (Sakamoto et al., 1992), as well as the gene structure and expression of CuZnSODs from rice were reported (Sakamoto et al., 1995a; Sakamoto et al., 1995b). In our laboratory, we are studying oxidative stress effect on the antioxidant enzymes of rice (Wei et al., 1995; Chen, 1994). In order to understand the regulation of rice SOD in response to a stress, cDNA clones of CuZnSOD and MnSOD were isolated and characterized from rice cDNA library (Pan et al., 1995; Chen et al., 1997). In this report, we describe the over-expression and characterization of CuZnSOD from rice in Escherichia coli, using the fusion protein to prepare antiserum. The antiserum that recognizes CuZnSOD was used to examine the SOD protein from different plant species.
1Corresponding author. Tel: 886-2-3630231 ext. 2366; Fax: 886-2-3918940; E-mail: pan@ccms.ntu.edu.tw