Bot. Bull. Acad. Sin. (1999) 40: 283-287
Chen and Kao Excess Cu and leaf senescence
Effect of excess copper on rice leaves: evidence for involvement of lipid peroxidation
Li-Men Chen and Ching-Huei Kao1
Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China
(Received August 11, 1998; Accepted April 30, 1999)
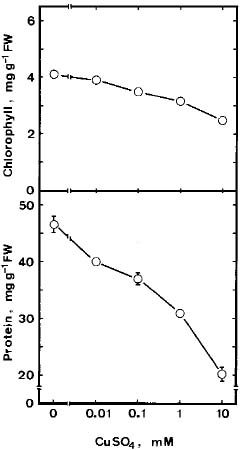
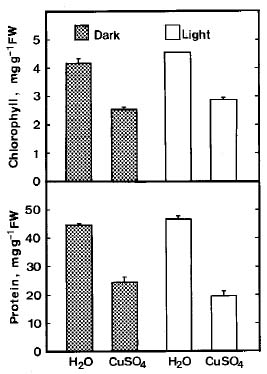
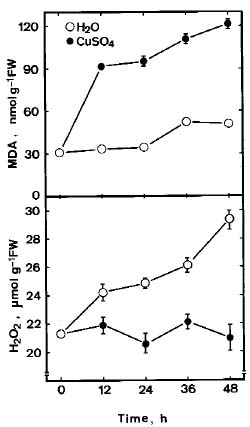
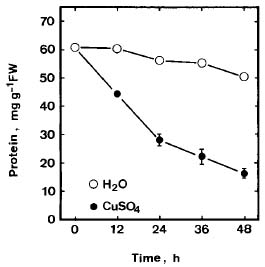
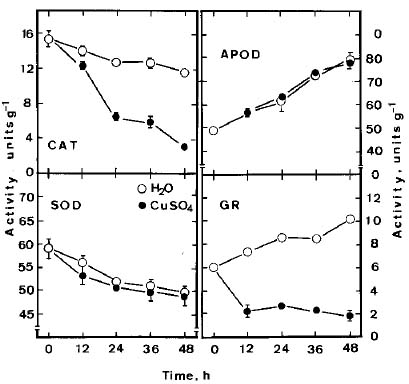
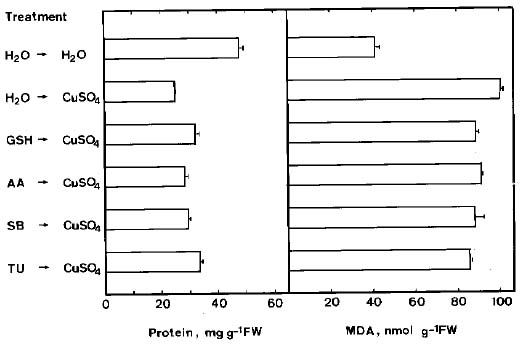
Abstract. Lipid peroxidation in relation to senescence of detached rice leaves caused by excess copper was investigated. Excess copper, which was found to promote senescence, increased the level of lipid peroxidation but not the level of H2O2. Catalase and glutathionine reductase activities were reduced by excess copper. Superoxide dismutase and ascobate peroxidase activities did not seem affected by excess copper. Free radical scavengers inhibited excess copper-promoted senescence and at the same time inhibited excess copper-induced lipid peroxidation, suggesting that lipid peroxidation induced by excess copper is mediated through free radicals.
Keywords: Copper; Lipid peroxidation; Leaf senescence; Oryza sativa.
Abbreviations: AA, ascorbic acid; APOD, ascobate peroxidase; CAT, catalase; GSH, reduced glutathione; GR, Glutathione reductase; MDA, malondialdehyde; SB, sodium benzoate; SOD, superoxide dismutase; TU, thiourea.
Introduction
Although Cu is an essential micronutrient for plants, uptake of excess Cu can be harmful to most plants (Fernandes and Henriques, 1991). It has been reported that Cu mediated free radical formation in isolated chloroplasts (Scandmann and Boger, 1980), in intact roots (De Vos et al., 1993), in detached leaves (Luna et al., 1994), and in intact leaves (Weckx and Clijsters, 1996). Free radical-induced lipid peroxidation is considered to be an important mechanism of leaf senescence (Dhindsa et al., 1981; Kunnert and Ederer, 1985; Slater, 1972; Strother, 1988; Thompson et al., 1987). Excess Cu has been shown to induce leaf senescence (Chen and Kao, 1998; Jana and Choudhuri, 1982; Luna et al., 1994). It appears that Cu-induced leaf senescence is associated with lipid peroxidation. In this study, effects of Cu excess on the senescence, lipid peroxidation, and on some enzymes of activated oxygen metabolism in detached rice leaves were investigated.
Materials and Methods
Rice (Oryza sativa cv. Taichung Native1) was cultured as previously described (Kao, 1980 ). The apical 3-cm segments excised from the third leaves of 12-day-old seedlings were used. Briefly, rice seedlings were planted on a stainless net floating on half-strength Johnson's modified nutrient solution (pH 4.2) in a 500 mL beaker. The nutrient solution was replaced every three days. Rice plants were grown for 12 days in a greenhouse, where natural
light was provided and the temperature was controlled at 30°C during the day and at 25°C at night. The apical 3 cm of the third leaf was used for the experiment. A group of 10 segments was floated in a Petri dish containing 10 mL of test solution. Incubation was carried out at 27°C in the light (40 µmol m-2 s-1) or in the dark.
Chlorophyll was determined according to Wintermans and De Mots (1965) after extraction in 96% (v/v) ethanol. For protein extraction, leaf segments were homogenized in 50 mM sodium phosphate buffer (pH 7.5). The extracts were centrifuged at 17,600 g for 20 min, and the supernatants were used for determination of protein by the method of Bradford (1976).
Malondialdehyde (MDA) was extracted with 5% (w/v) trichloroacetic acid and determined according to Heath and Packer (1968). MDA level is routinely used as an index of lipid peroxidation.
The H2O2 level was colorimetrically measured as described by Jana and Choudhuri (1981). H2O2 was extracted by homogenizing 50 mg leaf tissue with 3 mL of phosphate buffer (50 mM, pH 6.5). The homogenate was centrifuged at 6,000 g for 25 min. To determine H2O2 levels, 3 mL of extracted solution was mixed with 1 mL of 0.1% titanium sulfate in 20% (v/v) H2SO4. The mixture was then centrifuged at 6,000 g for 15 min. The intensity of the yellow colour of the supernatant at 410 nm was measured. H2O2 level was calculated using the extinction coefficient 0.28 µmol-1cm-1.
For extraction of enzymes, leaf tissues were homogenized with 0.1 M sodium phosphate buffer (pH 6.8) in a chilled pestle and mortar. The homogenate was centrifuged at 12,000 g for 20 min and the resulting supernatant was used for determination of enzyme activity. The whole
1Corresponding author. Fax: 886-2-2362-0879; E-mail: Kaoch@cc.ntu.edu.tw