Bot. Bull. Acad. Sin. (1999) 40: 295-304
Tzeng et al. Grapevine leafroll disease detection
Improvement of techniques for purification of leafroll
associated closterovirus from affected grapevines and the
preparation of antisera for disease indexing
Huey-Lin Chen Tzeng1, Moh-Jih Chen2, and Dean Der-Syh Tzeng2,3
1Department of Plant Pathology and Entomology, Taiwan Tobacco Research Institute, Tsao-Hu, Taichung 412, Taiwan
2Department of Plant Pathology, National Chung Hsing University, Taichung 402, Taiwan
(Received January 12, 1999; Accepted March 9, 1999)
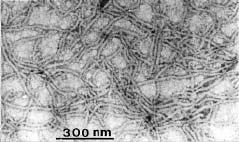
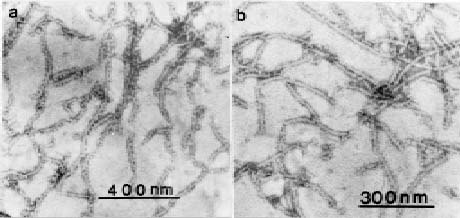
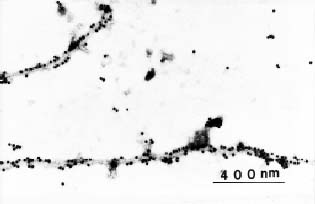
Abstract. An improved technique was developed for the purification of leafroll associated closterovirus (GLRaV) and successfully applied for the preparation of polyclonal and monoclonal antisera for the virus detection. The technique improvement mainly involves the use of a Chinese herb grinder for the homogenization of diseased sample. It requires very limited labor, is repeatable in regard to the virus yield, and can process considerable quantities within a short period. By this sample homogenization technique and a method modified from one in the available literature, GLRaV was successfully purified from diseased Kyoho, Black Queen, and Golden Mutscate grapevines cultivated in Taiwan. The electron microscopic examination of the obtained virus preparations revealed that the yield and purity were comparable or better than that reported by previous workers. With the obtained virus samples, polyclonal and monoclonal antisera were prepared. The results obtained from direct ELISA (enzyme linked immunosorbent assay) and indirect ELISA revealed that the prepared antisera detected mainly type III and type IV GLRaV. With the immunological method established, a disease survey was conducted between the years 1987 and 1994. A total of 204 grapevine cultivars or breeding lines were examined, and approximately 52% of them were found affected by GLRaV. Among the surveyed grapevines, about 30% were affected by Type III virus and another 31% by Type IV. A complexed infection of Types III and IV appeared to be common among Kyoho grapevines. The results also indicated the natural spread of GLRaV in tested field and suggested the need of a strict sanitation program for important breeding stock conservation.
Keywords: Antiserum preparation; Chinese herb grinder; Disease indexing; Grapevine leafroll disease; Virus purification.
Introduction
Leafroll is one of the most important diseases on cultivated grapevines and is a worldwide problem (Bovey et al., 1980). Research in recent years has indicated a consistent association of closterovirus particles with the leafroll affected grapevines. The etiological correlation of these closteroviruses and grapevine leafroll disease is now well accepted (Gugerli et al., 1984; Hu et al., 1989 & 1991; Zee et al., 1987; Zimmermann et al., 1988). Although it has been reported that mealybug (Pseudococcus spp.) (Engelbrecht and Kasdorf, 1990; Rosciglione and Gugerli, 1989; Tanne et al., 1989) and scale insect (Pulvinaria vitis L.) (Belli et al., 1994) may serve as agents for the disease transmission, it is generally believed that the widespread nature of this disease is primarily due to the use of diseased vines as propagating material. Strategies recommended for control of the disease in most grapevine growing countries mainly involves the use of virus free vines for the establishment or replacement of the vineyard.
The success of a clean seed program in the control of viral diseases depends greatly upon the technology and methodology available for the identification of the virus infection. The detection of GLRaV infection has traditionally been done solely by chip-bud grafting field indexing (Goheen, 1988). The main disadvantage of this methodology is that it requires a long process time and large amounts of field space and manpower. In addition, symptom development is often complicated by the presence of various biological and abiological stress factors (Egger et al., 1985; Ioannou and Gonsalves, 1991). Great improvement in the disease diagnosis was recently achieved by the successful development of virus specific antisera and the establishment of immunological detection techniques (Hu et al., 1989 and 1990a; Zee et al., 1987; Zimmermann et al., 1988 and 1990a,b). However, one of the major problems associated with the immunological detection of GLRaV is the limited availablity of antiserum; and what is even worse is the existence of at least seven different serotypes among the natural viral population on cultivated grapevines (Boscia et al., 1995; Choueiri et al., 1996; Hu et al., 1990a,b; Zimmermann et al., 1990a,b). In the Foundation Plant Material Service (FPMS) at the University of
3Corresponding author.