Bot. Bull. Acad. Sin. (1999) 40: 305-310
Hou et al. Fatty acid esterases from yam tuber
Purification and properties of fatty acid esterases from yam (Dioscorea batatas Decne) tuber
Wen-Chi Hou, Hsien-Jung Chen, Chung-Fang Chang and Yaw-Huei Lin1
Institute of Botany, Academia Sinica, Nankang, Taipei 115, Taiwan
(Received February 5, 1999; Accepted March 5, 1999)
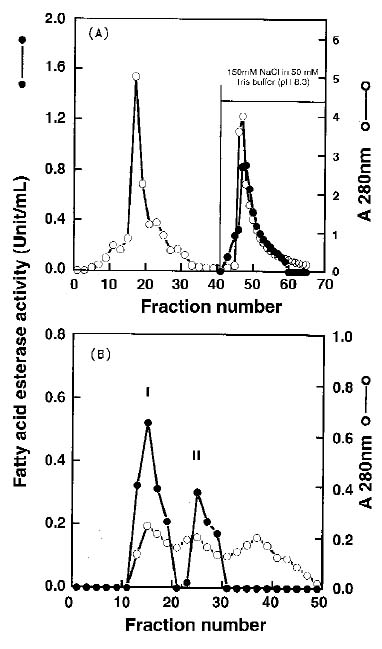
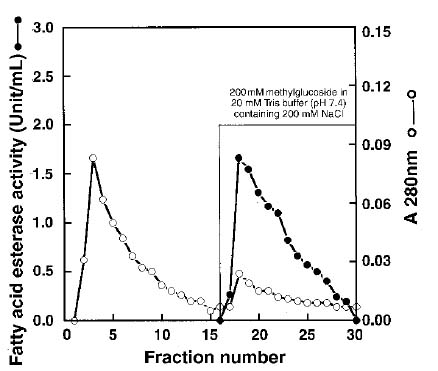
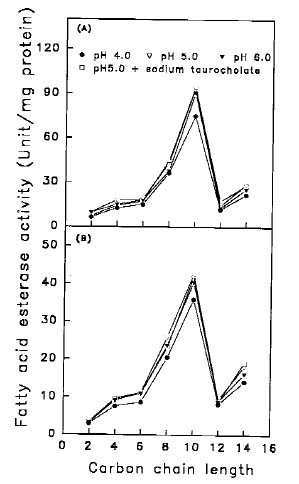
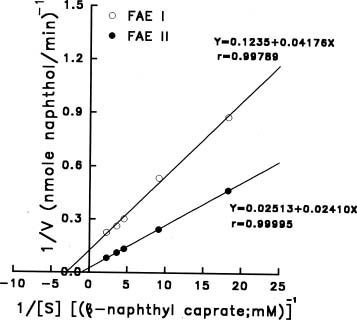
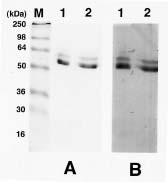
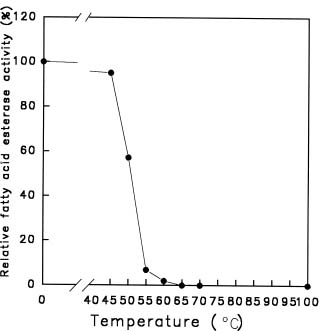
Abstract. Using b-naphthyl myristate (C14 fatty acid ester) as a screening substrate, we purified fatty acid esterases (FAEs) from yam (Dioscorea batatas Decne) tuber. Two FAE fractions (named FAE I and FAE II) were obtained after DE-52 ion exchange chromatography and Sephadex G-75 gel filtration purification steps, and then further purified by Con A Sepharose 4B affinity chromatography. FAE I and II fractions contained the same three protein bands about 50-64 kDa corresponding to esterase activity bands on SDS-PAGE gels. Among b-naphthyl esters determined at pH 4.0, 5.0 and 6.0 the best substrate for both FAE fractions was a C10-containing one with a maximum pH at 5.0. The Km and Vmax for b-naphthyl caprate (C10 fatty acid ester) of FAE I and II at 37°C, pH 5.0 were 0.338 and 0.959 mM; 0.405 and 0.585 nmole b-naphthol/min µg protein, respectively. FAE activity was stable below 50°C and lost completely above 65°C.
Keywords: Dioscorea batatas Decne; Fatty acid esterase (FAE); b-naphthyl caprate; b-naphthyl myristate; purification.
Abbreviations: APS, ammonium persulfate; Bis, N,N'-methylene-bis-acrylamide; CBB G-250, coomassie brilliant blue G-250; FAE, fatty acid esterase; 2-ME, 2-mercaptoethanol; SDS-PAGE, sodium dodecylsulfate-polyacrylamide gel electrophoresis; TCA, trichloroacetic acid; TEMED, N,N,N',N'-tetramethyl-ethylene-diamine.
Introduction
Multiple forms of esterase catalyzing the hydrolysis of carboxylic esters of short-chain fatty acids have been demonstrated in several plant tissues including leaves (Rudolph and Stahmann, 1966) and seeds (Schwartz et al., 1964; Veerabhadrappa and Montgomery, 1971) of Phaseolus species, carrot roots (Carino and Montgomery, 1968) and tubers of potato (Desborough and Peloquin, 1966; 1967; Galliard and Dennis, 1974). However, few works concerning long-chain fatty acid esters from plant sources have appeared except for studies of patatin, the potato tuber storage protein, which was demonstrated to have fatty acid esterase (Racusen, 1984), lipid acyl hydrolase, and acyltransferase activities involved wound responses of the tuber (Andrews et al., 1988; Jefferson et al., 1990).
Yam (Dioscorea species) is a member of the monocotyledonous family Dioscoreaceae and is a major tuber crop in West Africa, Southeast Asia and the Caribbean (Akoruda, 1984). We found yam (Dioscorea batatas Decne) tubers contained high fatty acid esterase (FAE) activities toward b -naphthyl esters of long-chain fatty acids in a preliminary work. In this report, we purified FAEs from yam tuber using b -naphthyl myristate as a screening substrate and characterized their properties.
Materials and Methods
Materials
Fresh tubers of yam (Dioscorea batatas Decne), imported from Japan, were purchased from a local market. Electrophoresis grade acrylamide and Bis, TEMED and APS were from E. Merck Inc. (Darmstadt, Germany). b -naphthyl esters of C2, C4, C6, C8, C10, C12 and C14 fatty acid were purchased from Sigma Chemical Company (St. Louis, MO, USA). A See BlueTM pre-stained standard kit for SDS-PAGE was obtained from NovexTM (San Diego, CA, USA). It contained myosin (250 kDa), BSA (98 kDa), glutamate dehydrogenase (64 kDa), alcohol dehydrogenase (50 kDa), carbonic anhydrase (36 kDa), myoglobin (30 kDa), and lysozyme (16 kDa). Con A Sepharose 4B and Sephadex G-75 gels were obtained from Pharmacia (Uppsala, Sweden).
Purification of FAEs from Yam Tuber
Extraction and purification processes were performed at 4°C. After being cleaned with water, yam tubers were peeled and cut into strips immediately for FAE extractions. Samples were homogenized with four volumes (W/V) of 50 mM Tris-HCl buffer (pH 8.3). After centrifugation at 14000 g for 30 min, the supernatants were saved and loaded onto a DE-52 ion exchange column (2.0 × 20 cm). The column was washed with 50 mM Tris-HCl buffer (pH 8.3) of three column volumes and then eluted batchwise with 150 mM NaCl in 50 mM Tris-HCl buffer (pH 8.3). Flow rate
1Corresponding author. Fax: 886-2-2782-7954; Tel: 886-2-2789-9590 ext. 320; E-mail: boyhlin@ccvax.sinica.edu.tw