Bot. Bull. Acad. Sin. (1999) 40: 311-317
Lee and Chung Genetic variation in Sasamorpha borealis
High levels of genetic variation in Korean populations of Sasamorpha borealis (Poaceae)
Nam Won Lee and Myong Gi Chung1
Department of Biology, Gyeongsang National University, Chinju 660-701, The Republic of Korea
(Received June 2, 1998; Accepted April 1, 1999)
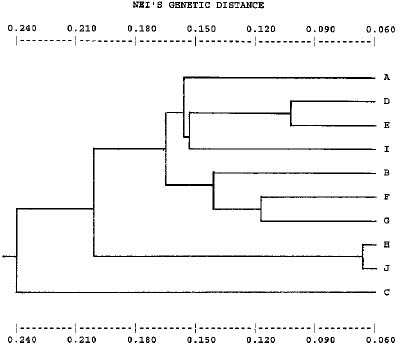
Abstract. Genetic and genotypic diversity of Korean populations in Sasamorpha borealis (Hack.) Nakai were investigated using allozymes as genetic markers. In Korea, S. borealis usually grows on hillsides under pine-oak understory. Many species of bamboos have the intermast period of "mast seeding." After spreading by extensive rhizomes for a specific period, nearly all adults in one area produce wind-pollinated flowers, set large quantities of seeds, and die. This study was undertaken to infer relationships between levels of genetic diversity and the reproductive biology of the species. Populations of S. borealis maintain high levels of genetic diversity: 51% of the loci examined were polymorphic and of mean genetic diversity (HeP = 0.219). The mean number of multilocus genotypes per population was 18.4, and the mean genotypic diversity index (DG) was 0.720. However, 31% of the total genetic variation was found among populations (GST = 0.310). Allele frequencies for all loci examined differed significantly among populations (p < 0.001). Average genetic identity for all pairs of populations was 0.833 (SD = 0.046). Indirect estimates of the number of migrants per generation (Nm) were 0.56 (calculated from GST) and 0.20 (calculated from the mean frequency of ten private alleles). Several biological and ecological traits found in S. borealis (widespread geographical distribution, long-lived habit, clonal reproduction with relatively high genotypic diversity, wind-pollinated breeding system, death of adults bearing large quantities of seeds, probable low gene flow among populations, selection for heterozyous genotypes in some of the sampled populations, the patterns of recolonization, effects of genetic drift, and local selective forces) may have played roles in shaping the population genetic structure of the species.
Keywords: Allozymes; Clonal diversity; Genetic diversity; Mast seeding; Sasamorpha borealis; Selection.
Introduction
Electrophoretic techniques for the study of clonal plants provide powerful genetic markers for recognizing individual plant genotypes (Berg and Hamrick, 1997). This approach has made it possible to better understand the spatial distributions of clones and the genotype diversity maintained within populations. According to a thorough review of studies of clonal plants (Ellstrand and Roose, 1987), species with predominantly vegetative reproduction generally have lower levels of genetic diversity than species that successfully produce progeny solely by sexual reproduction. Previous studies have revealed that vegetative reproduction and spread have a marked effect on the genetic structure of populations. As Eriksson (1993) pointed out, however, mechanisms for the maintenance of genetic variation in clonal plants are still controversial. For example, several results indicate that clonal reproduction could retard the loss of genetic diversity within populations because species with independent ramets could reduce the possibility of genet death (e.g., Parker and Hamrick, 1992; Berg and Hamrick, 1994; Lokker et al., 1994; Kim and Chung, 1995a,b; Mayers et al., 1998; Chung and Chung, 1999; Chung and Epperson, 1999). Others showed
that clonal reproduction may act as an enhancer of genetic drift by reducing the effective size of local populations (e.g., Pleasants and Wendel, 1989; Bayer, 1990; Murawski and Hamrick, 1990; Aspinwall and Christian, 1992; Chung, 1994; Chung and Kang, 1996; Yeeh et al., 1996). Since the relationship between genetic diversity and mode of reproduction remains unclear, further study of clonal plants is called for.
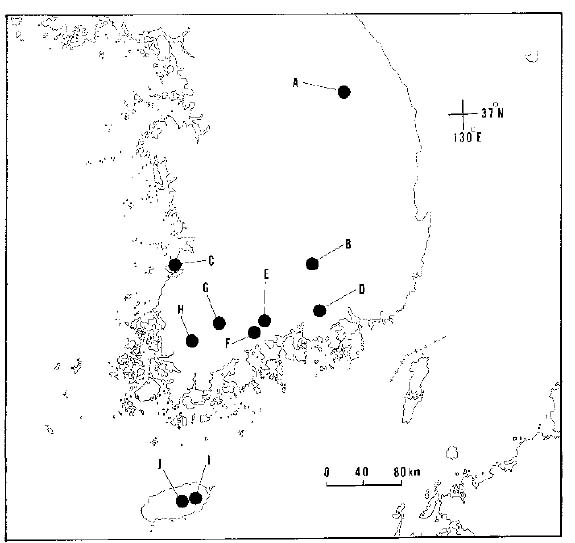
According to a previous review of bamboo biology (Janzen, 1976), many of the more common Asian species spread by rhizomes for a species-specific period (e.g., Sasa tessellata has an intermast period of > 115 years). After reaching a specific period, most Asian bamboos produce wind-pollinated flowers, synchronously set large quantities of seed ("mast seeding"), and die (Janzen, 1976). The biology of bamboos may be of interest to plant population biologists. Here we report levels and partitioning of allozyme diversity within and among populations and the extent of clonal reproduction within populations of Sasamorpha borealis (Hack.) Nakai (Poaceae), a small bamboo. Sasamorpha borealis is widely distributed in the Korean peninsula and Japanese archipelago. In Korea, populations of S. borealis are large and commonly found on hillsides under pine-oak understory. Unfortunately, information on reproductive biology of the study species (e.g., the exact period of masting and the range of synchronization of the masting period in Korea) is not available.
1Corresponding author. Fax: (0591) 54-0086; Tel: (0591) 751-5953; E-mail: mgchung@nongae.gsnu.ac.kr