Bot. Bull. Acad. Sin. (2000) 41: 19-25
Lee Acid phosphatase in Ulva
Phosphate starvation induction of acid phosphatase in Ulva lactuca L. (Ulvales, Chlorophyta)
Tse-Min Lee1
Institute of Marine Biology, National Sun Yat-sen University, Kaohsiung 80424, Taiwan
(Received November 23, 1998; Accepted March 5, 1999)
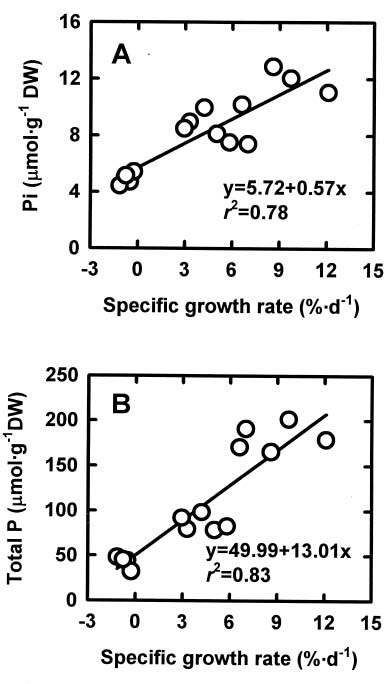
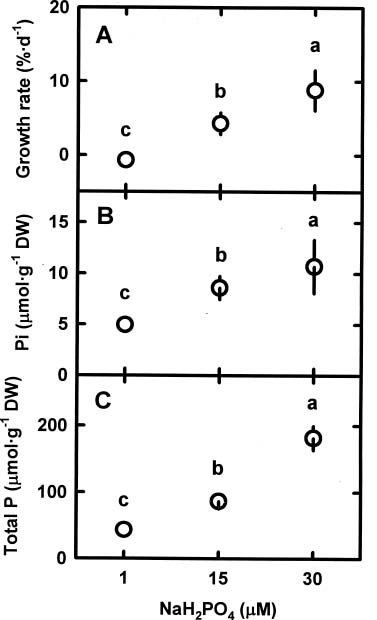
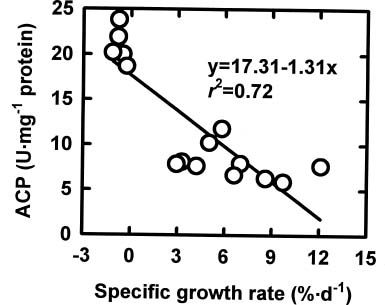
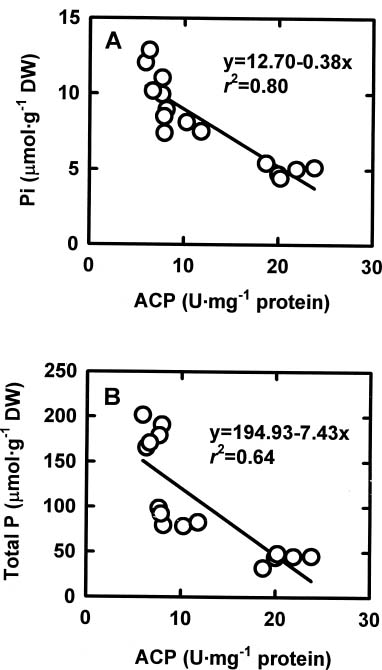
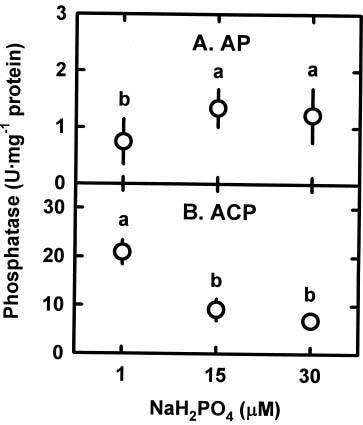
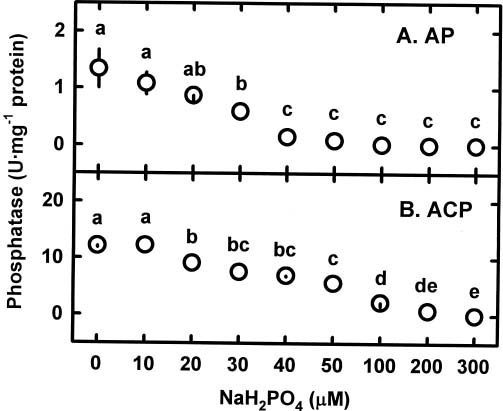
Abstract. The relationships among specific growth rate, intracellular inorganic phosphate (Pi) concentrations, total phosphorus (P) concentrations and specific activities of acid phosphatase (ACP; EC 3.1.3.2) and alkaline phosphatase (AP; EC 3.1.3.1) were studied in a green marine macroalga Ulva lactuca L. after 4 days of incubation at 1, 15 or 30 µM NaH2PO4. Both the specific growth rate and the intracellular Pi and total P concentrations increased with increasing NaH2PO4 levels; a linear correlation with specific growth rate was found for both intracellular Pi concentrations (r2=0.78, P<0.05) and the total P concentrations (r2=0.83, P<0.05). The growth rate and the Pi and total P concentrations were relatively low at 1 µM NaH2PO4, indicating U. lactuca L. could be P-deficient at 1 µM NaH2PO4. The specific activity of ACP was 4 to 10-fold greater than that of AP. Exposure to 1 µM NaH2PO4 increased the specific activity of ACP but slightly decreased that of AP. The ACP specific activity was inversely correlated with specific growth rate (r2=0.72, P<0.05) and also intracellular Pi concentrations (r2=0.80, P<0.05) and the total P concentrations (r2=0.64, P<0.05). The AP specific activity had no correlation with specific growth rate, intracellular Pi concentrations, or total P concentrations. The addition of NaH2PO4 concentrations higher than 20 µM to the assay mixture inhibited the specific activities of both ACP and AP. Overall, the P deficiency induction of ACP activity is related to a decrease in P availability in U. lactuca L.
Keywords: Acid phosphatase; Algae; Alkaline phosphatase; Phosphate starvation; Ulva lactuca.
Abbreviations: DW, dry weight; AP, alkaline phosphatase; ACP, acid phosphatase; P, phosphorus; Pi, inorganic oxyanion phosphate; p-NPP, p-nitrophenol phosphate.
Introduction
In algae, phosphorus (P) is an important and often limiting nutrient (Davies, 1988). P deficiency leads to metabolic disorders such as a decrease in photosynthesis and respiration, a retardation of growth, and even the death of algae cells (Davies, 1988; Gárciá-Sánchez et al., 1996; Lapointe, 1987; Theodorou et al., 1991). In general, algae take up P in the form of inorganic oxyanion phosphate (Pi) from the surrounding media via an active transport. However, most soluble Pi in seawater is esterified to organic compounds such as dissolved organic phosphomonoesters, rendering it is impossible for algae to uptake directly. It is known that most marine algae can synthesize extracellular alkaline phosphatase (AP; EC 3.1.3.1) to liberate Pi from dissolved organic phosphomonoesters, especially under P-deficient conditions (Fitzgerald and Nelson, 1966; Hernández et al., 1993; Weich and Graneli, 1989). In the case of the marine chlorophyte macroalga Ulva lactuca L., the activity of extracellular AP increased during cultivation in a P-deficient medium (Weich and Graneli, 1989). A similar trend was
also found in the freshwater microalgae Chlamydomonas reinhardtii grown in the absence of Pi (Quisel et al., 1996).
Algae can store P intracellularly as Pi or polyphosphate, which is utilized during the subsequent growth process or in P-deficient conditions (Cole and Huges, 1965; Kulaev and Vagabov, 1983; Lundberg et al., 1989; Watanabe et al., 1987). The concentrations of intracellular polyphosphates in U. lactuca L. decrease after transfer to P-deficient conditions, indicating that the intracellular polyphosphates could be hydrolyzed to meet P requirements (Weich and Graneli, 1989). Several intracellular phosphatases have been suggested to have a role in the hydrolysis of intracellular polyphosphates (Huber and Hamel, 1985; Marco and Orús, 1988). In microalgae, P-deficiency increases the activity of intracellular acid phosphatase (ACP; EC 3.1.3.2) (Blum, 1965; Price, 1962), which is localized in the cytoplasm (Moher et al., 1975; Price, 1962; Sommer and Blum, 1965). There are two kinds of ACP in microalgae: constitutive and inducible enzymes (Blum, 1965). In macroalgae, the responses of phosphatase to P-deficiency have been mainly studied on AP. Although the role of ACP in phosphorus metabolism has been extensively studied in terrestrial plants (Duff et al., 1994), we still have little knowledge about the relationships between intracellular soluble ACP and P availability in macroalgae.
1Tel: 886-7-5252000 ext. 5110; Fax: 886-7-5255100; E-mail: tmlee@mail.nsysu.edu.tw