Bot. Bull. Acad. Sin. (2000) 41: 27-34
Prado et al. Germination, growth and soluble sugar content
Effect of NaCl on germination, growth, and soluble sugar
content in Chenopodium quinoa Willd. seeds
Fernando E. Prado1,3, Cecilia Boero2, Miriam Gallardo2 and Juan A. González2
1 Cátedra de Fisiología Vegetal, Facultad de Ciencias Naturales e IML, Miguel Lillo 205, CP 4000 Tucumán, Argentina
2 Laboratorio de Botánica, Fundación Miguel Lillo, Miguel Lillo 251, CP 4000, Tucumán, Argentina
(Received December 22, 1998; Accepted May 18, 1999)
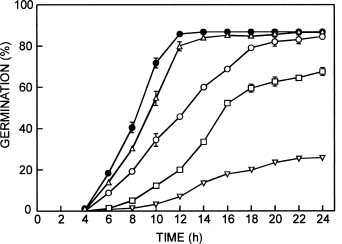
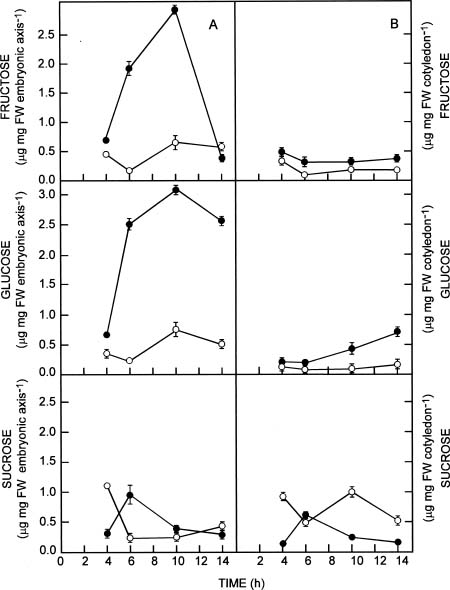
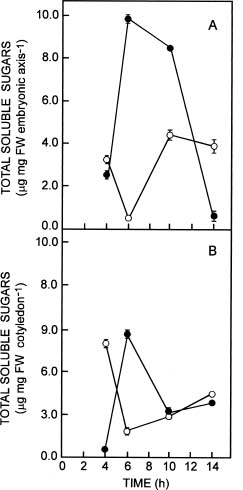
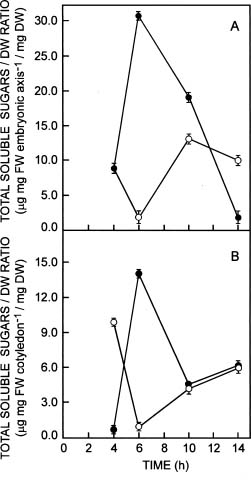
Abstract. The influence of NaCl on germination rate, growth, and soluble sugar content in quinoa (Chenopodium quinoa Willd.) seeds and seedling components (cotyledons and embryonic axes) during early germination was investigated. Under saline conditions germination decreased markedly. The delay in germination rate was not linear. In presence of 0.4 M NaCl, the percentage of germination was only 14% after 14 h, whereas the control at the same time reached maximum germination (87%). The percentage of aborted seeds after 14 h in NaCl was lower than in distilled water (7% and 16%, respectively). A high percentage (67%) of the ungerminated seeds from saline treatment germinated after washing with distilled water. Fresh weight was reduced by salinity in both embryonic axes and cotyledons. The highest increase in fresh weight was observed in embryonic axes, where an increase up to 3-fold higher occurred in distilled water than in saline conditions after 10 h of development. The water content of embryonic axes in distilled water increased considerably for the first 10 h, then remained constant thereafter. In saline conditions, no significant changes were observed in water content. NaCl did not strongly inhibit dry mass production in either embryonic axes or cotyledons. Total soluble sugar content increased markedly in distilled water, peaking after 6 h for both embryonic axes and cotyledons. Reduced glucose and fructose contents were found in embryonic axes in the presence of NaCl. However, in the cotyledons, the glucose and fructose contents did not differ significantly. Levels of sucrose were higher in NaCl-treated cotyledons than in control seeds. The relationships between germination, water content, seedling growth, and soluble sugar content in relation to salt stress are discussed.
Keywords: Chenopodium quinoa; Germination; Growth; Saline stress; Seeds; Soluble sugar content.
Introduction
Quinoa (Chenopodium quinoa Willd.) is a dicotyledoneous species native to the Andean Region of South America, where it is used as a very important nutritional resource (Risi and Galwey, 1984). This species is among the most important grain crops in terms of protein content and amino acid balance for human nutrition because of its high lysine and methionine levels (González et al., 1989; Burnouf-Radosevich, 1988; Jacobsen, 1993). A large spectrum of nutritious and other useful products is being made from quinoa for human or animal consumption (Jacobsen and Stølen, 1993). Quinoa and other halophytic members of the Chenopodiaceae have long been known for their extraordinary salt and drought tolerances (Reimann and Breckle, 1993). These characteristics make it an attractive alternative crop for the arid and semiarid regions, where water deficiency and salinity have been recognized as major agricultural problems (Aronson, 1985). In halophytes adaptation to salinity is associated with metabolic adjustments that lead to the accumulation of several organic solutes, such as sugars, polyols, betaines,
and free proline (Flowers et al., 1977; Briens and Larher, 1982; Gorham et al., 1981).
The sensitivity of plants to salinity may depend on their developmental stage (Adam, 1990). Different responses to salinity have been reported between germinating and growing seedlings of a number of halophytes (Mayer and Poljakoff-Mayber, 1975). Ungar (1996) observed that seeds of Atriplex patula were less affected by salinity than the growing plants. Nevertheless, very little information is available regarding the relative salt tolerance of plants at different stages of development (Ungar, 1991). The results obtained to date are not clear, even though plant response to salinity has been one of the most widely researched subjects (Munns, 1993).
Carbohydrates, especially starch, represent the major reserve substance in most seeds (Bewley and Black, 1994). During early germination, mobilisation of storage carbohydrates occurs, especially after radicle emergence. However, in the growing regions, i.e., embryonic axes, some mobilisation can occur before germination is completed (Bewley and Black, 1994). Once the high molecular weight carbohydrates are mobilised, they are converted into soluble forms, i.e., sucrose, glucose and fructose, that are readily transportable to sites where they are required for growth (Mayer and Poljakoff-Mayber, 1975). The
3Corresponding author. Fax: 0054-081-239456 or 0054-081-330868; E-mail: decano@csnat.unt.edu.ar or fepra@unt.edu.ar