Bot. Bull. Acad. Sin. (2000) 41: 41-48
Jang et al. PUFA production by solid substrate fermentation
Polyunsaturated fatty acid production with Mortierella alpina by solid substrate fermentation
Hung-Der Jang, Yuh-Yih Lin and Shang-Shyng Yang1
Department of Agricultural Chemistry, National Taiwan University, Taipei 10617, Taiwan
(Received April 16, 1999; Accepted May 28, 1999)
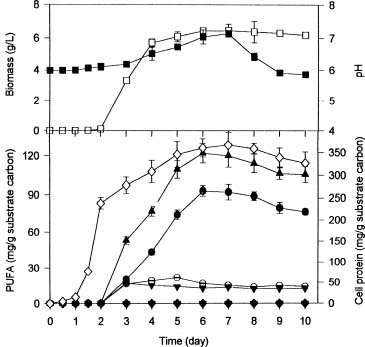
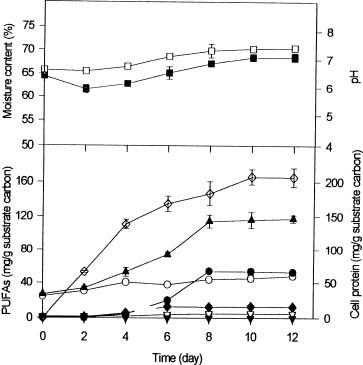
Abstract. Polyunsaturated fatty acids (PUFA) were produced with Mortierella alpina by solid substrate fermentation. Rice bran was the most effective substrate for PUFA production, followed by peanut meal residue, wheat bran, and sweet potato residue. The optimal conditions for PUFA production were rice bran supplemented with 2.3 to 5% nitrogen at an initial moisture content of 65 to 68% and a pH range of 6 to 7. Each gram of substrate carbon yielded 122.2 mg of total PUFA, including 12.8 mg of eicosapentaenoic acid (EPA), 47.8 mg of linoleic acid (LA), 7.1 mg of a-linolenic acid (ALA), and 54.5 mg of arachidonic acid (ARA) for 8 to 12 days incubation. C/N ratios between 14.5 and 18.5 favored EPA and LA production, while C/N ratios between 19.8 and 21 enhanced ARA and total PUFA production. Total PUFA, EPA and ARA production increased 12, 84.4 and 46.1%, respectively, when the culture temperature was shifted from 20°C to 12°C on the fifth day. Supplement of soybean and linseed oils increased LA by 84.9 and 36%, ARA by 71 and 42.1%, and EPA by 130.6 and 92.1%, respectively.
Keywords: Mortierella alpina; Polyunsaturated fatty acid; Rice bran; Solid substrate fermentation.
Introduction
The w-3 and w-6 series of polyunsaturated fatty acids have shown tremendous potential for use in food additives and pharmaceuticals for heart and circulatory disorders and cancer as well as inflammatory diseases (Dyerberg, 1986; Reddy and Maruyama, 1986). The submerged culture of Mortierella is usually used for PUFA production with glucose or glycerol as a carbon source (Bajpai et al., 1991; Li and Ward, 1994). However, submerged culture needs a higher energy input and produces more wastewater (Canpel and Moo-Young, 1980; Yang, 1988). In addition, culture media and culture conditions affect the quality and the quantity of PUFA production (Granger et al., 1992; Li and Ward, 1994).
Agricultural wastessuch as rice bran, wheat bran, peanut meal residue, and sweet potato residueare abundant in Taiwan. Both rice bran and wheat bran are good substrates for enzyme and oil production (Deschamps et al., 1985; Yang and Chiu, 1986; Yang and Wang, 1999). Being economically competitive, rice bran was used as the basal substrate in this report to produce PUFA with Mortierella alpina by solid substrate fermentation, and the optimal conditions for PUFA production were also investigated.
Materials and Methods
Microorganism
Mortierella alpina ATCC 32222 was purchased from American Type Culture Collection and used for the production of polyunsaturated fatty acids.
Solid Substrate
Rice bran, wheat bran, peanut meal residue, and sweet potato residue were purchased in a local market. The chemical composition of the test solid substrate is listed in Table 1.
Culture Media and Conditions
Mortierella was grown at 20°C in a membrane culture containing (mg l-1): glucose, 10; yeast extract, 5 and agar, 20 at pH 6.5. Mycelia were harvested from membrane culture and blended with a micro-Waring blender for mycelial suspension.
Submerged and Solid State Fermentation
Submerged basal medium contained (mg l-1) soluble starch, 20; Bacto yeast extract, 5; KNO3, 10; KH2PO4, 1 and MgSO47H2O, 0.5 at pH 6.5. The broth was inoculated with 5% (v/v) mycelial suspension and shaken at 200 rev min-1 and at 20°C for 2 to 10 days. Each ml of mycelial suspension contained 1.0-1.5 × 106 mycelial fragments.
1Corresponding author. Tel: 886-2-23621519; Fax: 886-2-23679827; E-mail: ssyang@ccms.ntu.edu.tw