Bot. Bull. Acad. Sin. (2000) 41: 57-60
Zheng and Ho The sexual stage of Phytophthora polygoni
The sexual stage of Phytophthora polygoni Saw.
X.B. Zheng1 and H.H. Ho2,3
1Plant Pathogen Research Institute, Nanjing Agricultural University, Nanjing 210095, Peoples Republic of China
2Department of Biology, State University of New York, New Paltz, New York 12561, USA
(Received March 1, 1999; Accepted May 20, 1999)
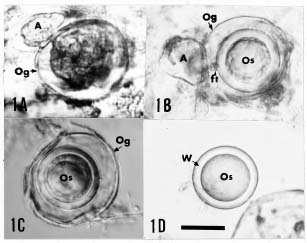
Abstract. The sexual stage of Phytophthora polygoni Saw.which is distinguishable from other species of Phytophthora by its inability to grow on common agar media, its sporangial characteristics and its disease symptomshas been successfully induced to form sexual organs in diseased leaves collected from the field or by artificial inoculation. The oogonia are smooth, spherical, thin-walled, (28.5-) 33 (-42) µm diam, with an aplerotic to markedly aplerotic, thick walled (3-4 µm) oospore, (24.7-) 28.1 (-32.3) µm diam, and a paragynous antheridium, 13-25 × 10-15 µm, attached laterally or apically to the oogonium. Interesting chlamydospore-like bodies were also observed.
Keywords: China; Phytophthora polygoni; Rumex dentatus; Sex organs.
Introduction
Phytophthora polygoni Saw., first described as a new species by Sawada (1922) causing leaf spot disease of Polygonium japonicum Meisn. in Taiwan was recently rediscovered on Rumex dendatus L. in Nanjing in Jiangsu Province of mainland China. It was identified based on similar disease symptoms and sporangial characteristics (Zheng and Ho, 1998). However, the fungus could not be isolated on culture media, and the sexual stage had not been found. Consequently, the species was excluded from treatment of the genus Phytophthora (Waterhouse, 1963) due to incomplete knowledge of its life cycle. Reported here are successful attempts to induce, under laboratory conditions, the formation of sex organs on diseased leaves of R. dentatus collected from the field and by artificial inoculation of healthy leaves with P. polygoni.
Materials and Methods
Diseased leaves of R. dentatus infected by P. polygoni were collected from Nanjing (late April to early July, 1998) and Yangzhou (early June, 1998) in Chinas Jiangsu Province. Freshly collected leaves (2-3) were placed in 12-15 ml sterile tap water in a 9-cm petri dish. Three replicates were used for each collection from the same locality at a specific time. The dishes were incubated in darkness for 12-15 days at 25, 20 and 14°C. At the end of the incubation period, a small piece of rotten tissue was placed on a glass slide. The excess water was removed with filter paper. The leaf tissues were cut into smaller pieces with a scalpel. Lactophenol (2-3 drops) was added to the specimen, heated over an alcohol lamp until it just started to boil, then removed from the flame until the boiling
stopped before it was reheated again. The procedure was repeated 3-5 times so that the tissues became clarified. They were then covered with a cover glass for studies under a light microscope.
For artificial inoculation, freshly collected diseased leaves were first rinsed clean under running tap water. Small pieces of leaf lesions (ca. 15 × 15 mm) were cut out and placed in 10 ml sterile tap water in a 9-cm petri dish, with the leaf surface facing upwards. After incubation under light at 20°C, abundant zoospores were released into the water. The zoospore suspension was transferred to a misting gun and sprayed on the healthy leaves of potted R. dentatus from the greenhouse. Then the entire plant was wrapped in a plastic bag for 24 h to maintain saturated humidity. At the end, the treated plants were kept in a greenhouse (25-28°C) for 5-7 days. When leaf lesions developed fully, diseased leaves were submerged in water and heated with lactophenol as described above.
Results
Sexual structures that were not found at any stage in the diseased leaves in the field, could be induced to produce in the laboratory by the methods described (Figure 1A-D). The sexual organs were frequently formed along and within the leaf veins and most abundantly in the midrib and petiole (adjacent to the lamina) of the diseased leaf. The mature oogoonia (Figure 1B-C) were smooth, spherical, thin-walled, straw-colored, measuring (28.5-) 33 (-42) µm diam with an aplerotic to markedly aplerotic spherical oospore (24.7-) 28.1 (-32.3) µm diam. The single-layered oospore wall was smooth and uniformly thick (3-4 µm) (Figure 1D). The paragynous antheridium 13-25 × 10-15 µm was attached laterally or apically to the side of the oogonium (Figure 1A-B). Presumably, fertilization occurred when the antheridial contents were discharged into the oogonium through a fertilization tube.
3Corresponding author. Tel: 914-257-3780; Fax: 914-257-3791; E-mail: hoh@matrix.newpaltz.edu