Bot. Bull. Acad. Sin. (2000) 41: 85-92
Chiang and Schaal Evolution of a cpDNA noncoding spacer
Molecular evolution of the atpB-rbcL noncoding spacer of
chloroplast DNA in the moss family Hylocomiaceae
Tzen-Yuh Chiang1,3 and Barbara A. Schaal2
1Department of Biology, Cheng-Kung University, Tainan 700, Taiwan
2Department of Biology, Washington University, St. Louis, MO 63130, USA
(Received January 20, 1999; Accepted July 28, 1999)
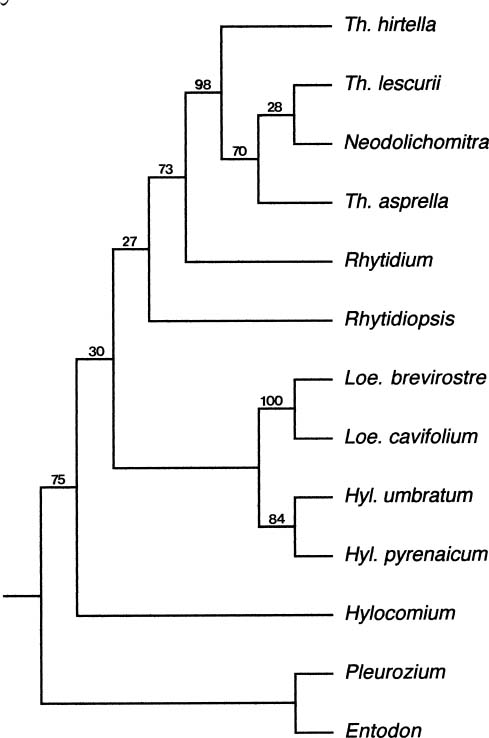
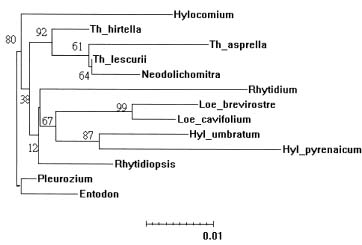
Abstract. Molecular evolution of the chloroplast noncoding region between the atpB and rbcL genes was investigated in the moss family Hylocomiaceae. Nucleotide substitution contributed to most of the variation among taxa, although an insertion of 29 base pairs was found in Rhytidiopsis robusta. The evolution of atpB-rbcL intergenic spacer was constrained in Hylocomiaceae compared to the evolutionary rates of this chloroplast spacer, and even of the rbcL gene, in angiosperms. Using a relative rate test procedure, substitution rates of the chloroplast spacer were found to be consistent with the hypothesis of a molecular clock, except for three taxa. Based on previous knowledge of the evolutionary rate of this spacer in true mosses, the divergence of the Hylocomiaceae from their common ancestor was estimated to be 29.0 million years ago, which is consistent with the fossil record. The chloroplast sequences supported the monophyly of the Hylocomiaceae with a bootstrap value of 82%. The effects of long branch attraction caused the erroneous placements of Rhytidiopsis and Rhytidium in the parsimony analysis. In contrast, neighbor-joining analysis provided a more congruent estimate of the phylogeny of the Hylocomiaceae based on the cpDNA variation observed.
Keywords: atpB-rbcL intergenic spacer; Chloroplast DNA; DNA sequence; Gene tree; Hylocomiaceae; Long branch attraction; Molecular clock; Mosses.
Introduction
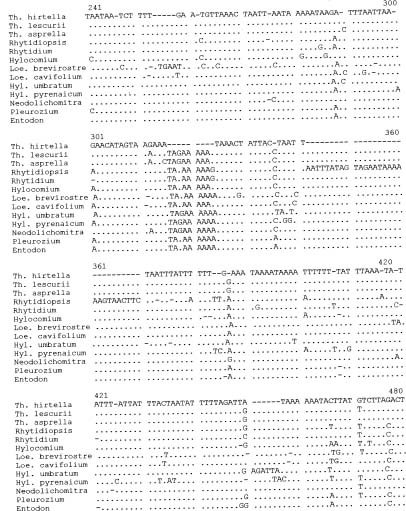
The atpB-rbcL spacer, a noncoding region of the chloroplast genome, has been recently used in phylogenetic studies of angiosperms (Golenberg et al., 1993; Manen et al., 1994b), such as the Betulaceae (Bousquet et al., 1992a), Celastrales (Savolainen et al., 1994) and Rubiaceae (Manen et al., 1994a; Manen and Natali, 1995; Natali et al., 1995), as well as of mosses (Chiang, 1994). The length of the spacer region varies from 519 to 557 base pairs in true mosses, which is longer than in the liverwort Marchantia polymorpha. The evolution of the atpB-rbcL spacer sequence of the true mosses (Chiang, 1994) is constrained relative to the spacer in angiosperms as well as to the rbcL gene in angiosperms (Zurawski et al., 1984; Manen and Natali, 1995).
In a previous study, the evolution of the chloroplast atpB-rbcL spacer in 11 families of true mosses was analyzed (Chiang, 1994). At this broad level, the spacer evolved at a rate consistent with the hypothesis of a molecular clock. In this study, we investigated the tempo and mode in evolution of the atpB-rbcL chloroplast spacer within the family Hylocomiaceae, which are widespread in temperate regions and occur on high mountains in the tropics (Rohrer, 1985). The phylogenetic relationships among
genera in this family have been determined based on the sequences of both the internal transcribed spacers of nuclear ribosomal DNA and the atpB-rbcL chloroplast spacer (Chiang, 1994). In contrast to a cladistic analysis based on morphological traits of the Hylocomiaceae (Rohrer, 1985), which recognized 12 genera, our study excluded six genera [i.e. Pleurozium, Rhytidiadelphus, Macrothamnium, Orontobryum, Leptohymenium, and Leptocladiella (cf. Chiang, 1995)] from the family. In addition, Thelia, a genus endemic to North America, was included as a member of the Hylocomiaceae based on molecular evidence (Chiang, 1994), bringing the genus number of the family up to seven.
This study had four goals: 1) to reconstruct the gene tree of the atpB-rbcL spacer in the family Hylocomiaceae; 2) to investigate the evolutionary rates of this spacer; 3) to test the hypothesis of a molecular clock; and 4) to estimate the time of divergence from a common ancestor of the Hylocomiaceae.
Materials and Methods
Plant Materials
According to a previously inferred phylogeny, eleven species of seven genera have been included in the Hylocomiaceae (Chiang, 1994). For the current study, all species of the Hylocomiaceae as well as two outgroup taxa, Pleurozium schreberi (Brid.) Mitt. and Entodon seductrix (Hedw.) C. Muell., were included (Table 1)
3Corresponding author. Tel: +886-6-2757575 ext. 65525; Fax: +886-6-2742583; E-mail: tychiang@ccvax.sinica.edu.tw