Bot. Bull. Acad. Sin. (2000) 41: 93-98
Chen et al. RP3 promoter expression and pigment contents
Expression of rice prolamin RP3 promoter exhibits a positive association with cellular pigment contents in transgenic tobacco suspension cultures
Hsien-Jung Chen1, Chih-Yuan Yang1, Yean-Ching Chen1 and Ching-San Chen1,2
1Institute of Botany, Academia Sinica, Nankang, Taipei, Taiwan 11529, Republic of China
(Received March 3, 1999; Accepted September 23, 1999)
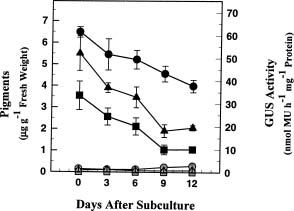
Abstract. Three transgenic tobacco regenerants (lines RP3/2, RP3/7 and RP3/18) harboring rice RP3/GUS chimeric gene inserts in their genomes were isolated previously and used to establish cell suspension cultures (Yang et al., 2000). A differential GUS expression pattern together with morphological, biochemical, and molecular variations were observed among these cell lines (Chen et al., 1999). In this report we used pigments such as carotenoids and chlorophylls a and b as markers to study the possible relationship between RP3 promoter activity and cellular pigment contents, and a positive association was found among these cell lines. RP3/2, which contained the highest cellular pigment contents, also exhibited the highest GUS expression level. The RP3 promoter activity in RP3/2 continuously decreased and was parallel with the reduction of pigment contents within a 12-day growth period after subculture. In RP3/18, both also showed a parallel association and remained relatively constant during cell growth. Based on these results, a positive association between RP3 promoter activity and cellular pigment content was found among and within cell lines. The meaning of the association and its possible explanation are discussed.
Keywords: Pigment; Prolamin; Rice; RP3 promoter; Tobacco.
Introduction
Rice is an important cereal crop in Asia and contains prolamins and glutelins as its major seed storage proteins (Bietz, 1982; Juliano, 1972). Prolamins form a small gene family (Kim and Okita, 1988a) and are comprised of heterogeneous proteins (Hibino et al., 1989) with molecular weights of 12 to 17 kDa (Mandac and Juliano, 1978; Padhye and Salunkhe, 1979). Prolamins constitute ca. 6 to 25% of total seed storage proteins on a weight basis, depending on the genotypes (Huebner et al., 1990), subcellular fractionation and pepsin digestion assay (Ogawa et al., 1987), and different extraction solvents (Sugimoto et al., 1986). Expression of rice prolamin is seed-specific and can be detected as early as 5 to 8 days after flowering by Northern blot (Kim et al., 1993; Shyur et al., 1992) or as early as 8 to 11 days using Western blot (Li and Okita, 1993; Shyur et al., 1994). Prolamins accumulate within protein body-I formed by direct dilation of the endoplasmic reticulum membrane (Krishnan et al., 1986; Yamagata and Tanaka, 1986).
Several rice prolamin genes have been isolated previously (Barbier and Ishihama, 1990; Feng et al., 1990; Kim and Okita, 1988a & b; Masumura et al., 1990; Shyur and Chen, 1993; Shyur et al., 1992; Wen et al., 1993; Yamagata
et al., 1992). However, factors involved in the prolamin gene expression and regulation are not yet clear. In our laboratory three genomic DNA clones (RP3, RP5 and RP6) of rice prolamins were isolated (Chen et al., 1996; Shyur et al., 1992; Wen et al., 1993). The RP3 promoter has been constructed with a bacterial GUS reporter gene and expressed in the embryo and endosperm tissues of transgenic tobacco seeds, and in the suspension-cultured cells established from transgenic tobacco regenerants (Yang et al., 2000). Characterization of these tobacco suspension cultures derived from transgenic regenerants showed that the line RP3/2 exhibited distinct sub-organelle structures of chloroplasts, and over-expressed the pigment contents and mRNA of a 23 kDa polypeptide of PSII oxygen-evolving complex (Chen et al., 1999). In this report a positive association between the GUS expression level and cellular pigment contents was observed. The indication and possible explanation for the association are discussed.
Materials and Methods
Plant Materials
The rice RP3 gene was isolated previously and encodes a putative seed prolamin storage protein (Chen et al., 1996). Transgenic tobacco regenerants (lines RP3/2, RP3/7 and RP3/18) harboring rice RP3/GUS chimeric gene construct in their genomes were isolated previously (Yang et al., 2000) and used to establish cell suspension cultures (Chen et al., 1999).
2Corresponding author. Tel: 886-2-27899590 ext. 251; Fax: 886-2-27858936; E-mail: bocschen@ccvax.sinica.edu.tw