Bot. Bull. Acad. Sin. (2000) 41: 99-103
Chen et al. Oxidative stress in copper-treated rice roots
Copper toxicity in rice seedlings: Changes in antioxidative enzyme activities, H2O2 level, and cell wall peroxidase activity in roots
Li-Men Chen, Chuan Chi Lin, and Ching Huei Kao1
Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China
(Received March 15, 1999; Accepted September 2, 1999)
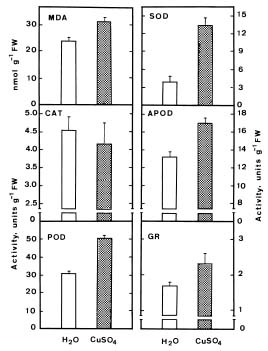
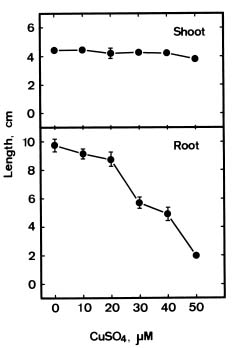
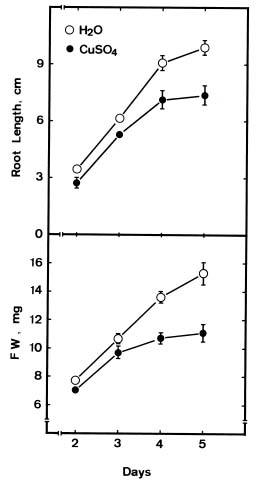
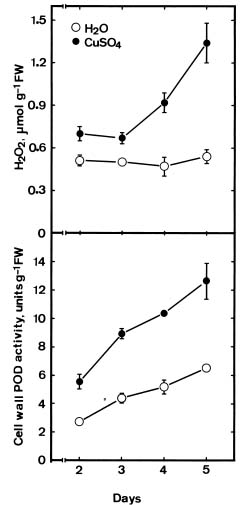
Abstract. The changes in lipid peroxidation, antioxidative enzyme activity, H2O2 level, and cell wall peroxidase activity in Cu-stressed roots of rice seedlings and their relation with root growth inhibition were investigated. CuSO4 was effective in inhibiting root growth but not shoot growth. Treatment with CuSO4 resulted in an increase in lipid peroxidation and modulated antioxidative enzyme activities in rice roots. CuSO4 increased the activities of superoxide dismutase, ascorbate peroxidase, glutathione reductase, and peroxidase, but had no effect on catalase. CuSO4 also increased H2O2 level and cell wall peroxidase in roots of rice seedlings. Exogenous application of H2O2 resulted in an inhibition of root growth. It appears that growth inhibition of root caused by Cu is associated with H2O2 dependent peroxidase-catalyzed formation of cross-linking among cell wall polymers.
Keywords: Copper; Lipid peroxidation; Oryza sativa; Oxidative stress; Root growth.
Abbreviations: APOD, ascorbate peroxidase; CAT, catalase; FW, fresh weight; GR, glutathione reductase; MDA, malondialdehyde; POD, peroxidase; SOD, superoxide dismutase.
Introduction
Copper (Cu) is an essential element for plant growth (Arnon and Stout, 1939), and it is important in various biochemical processes, but at toxic concentrations it interferes with numerous physiological processes (Fernandes and Henriques, 1991).
Cu is known to damage cell membranes by binding to the sulphydryl groups of membrane proteins and by inducing lipid peroxidation (De Vos et al., 1989; De Vos et al., 1992). Cu-mediated free radical formation has been demonstrated in isolated chloroplasts (Sandmann and Boger, 1980), in intact roots (De Vos et al., 1993), in leaf segments (Chen and Kao, 1999; Gallego et al., 1996; Luna et al., 1994) and in intact leaves (Weckx and Clijsters, 1996).
Cellular damage caused by free radicals might be reduced or prevented by a protective metabolism involving antioxidative enzymes such as SOD, APOD, GR, CAT and POD. SOD catalyzes the dismutation of two molecules of superoxide into oxygen and H2O2. APOD reduces H2O2 to water, with ascorbate as electron donor (Asada, 1992). GR plays a part in the control of endogenous H2O2 through an oxido-reduction cycle involving glutathione and ascorbate (Foyer and Halliwell, 1976; Smith et al., 1989). CAT and POD are implicated in removal of H2O2. It has been
reported that Cu increases the activities of antioxidative enzymes such as SOD (Chongpraditnum et al., 1992; Rama Devi and Prasad, 1998), POD (Karataglis et al., 1991), CAT (Rama Devi and Prasad, 1998), and APOD (Rama Devi and Prasad, 1998). It is well known that CAT and APOD play an important role in preventing oxidative stress by catalyzing the reduction of H2O2 (Weckx and Clijsters, 1996). Rama Devi and Prasad (1998) found that CAT and APOD activities were increased by Cu, suggesting that excess Cu may increase the production of H2O2. H2O2 is a necessary substrate for the cell wall stiffening process catalyzed by cell wall POD (Elstner and Heupel, 1976; Hohl et al., 1995; Schopfer, 1996), which is considered to be one of the mechanisms resulting in growth inhibition (Fry, 1986). The present investigation was designed to study the change in lipid peroxidation, antioxidative enzyme activities, H2O2 level and cell wall POD activity in Cu-stressed roots of rice seedling and their relation with root growth inhibition.
Materials and Methods
Rice (Oryza sativa L. cv. Taichung Native 1) seeds were sterilized with 2.5% sodium hypochlorite for 15 min and washed thoroughly with distilled water. These seeds were then germinated in a Petri dish (20 cm) containing distilled water at 37°C in the dark. After a 1-day incubation, uniformly germinated seeds were selected and transferred to Petri dishes (9.0 cm) containing two sheets of Whatman No. 1 filter paper moistened with 10 ml of distilled water
1Corresponding author. Fax: 886-2-23620879; E-mail: kaoch@ccms.ntu.edu.tw