Bot. Bull. Acad. Sin. (2000) 41: 129-137
Yang and Yang Xanthomonas campestris pv. citri recA gene
Construction and characterization of a recA mutant of Xanthomonas campestris pv. citri
Yen-Chun Yang and Mei-Kwei Yang1
Department of Biology, Fu Jen University, 510 Chun-Chen Road, Taipei 242, Taiwan, Republic of China
(Received August 23, 1999; Accepted October 25, 1999)
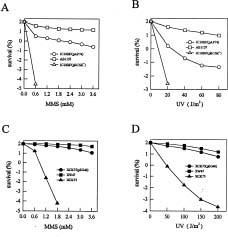
Abstract. The recA gene of Xanthomonas campestris pathovar citri (X. c. pv. citri) was cloned and sequenced. Nucleotide sequence analysis revealed an open reading frame of 1,032 bp that encodes a 344-amino acid protein and which shows a high degree of homology to recA genes from other bacteria. The cloned recA gene of X. c. pv. citri restored to a recA deletion mutant of Escherichia coli the ability to resist killing induced by methylmethane sulfonate or ultraviolet radiation, indicating that the cloned gene was functional in E. coli. A recA mutant of X. c. pv. citri was constructed by gene replacement and was shown both not to possess detectable DNA recombination activity and to be markedly more sensitive to DNA-damaging agents than is wild-type X. c. pv. citri. Transformation of recA mutants of E. coli and X. c. pv. citri with a plasmid containing X. c. pv. citri recA conferred the ability to produce a 37-kDa protein that reacted with antiserum to E. coli RecA protein. Two additional open reading frames were also identified in the recA region of the X. c. pv. citri genome: One located immediately upstream from recA that encodes a 213-amino acid protein with a high degree of similarity to the LexA protein of E. coli, and another located immediately downstream of recA that encodes a 153-amino acid protein with sequence similarity to the RecX protein of Pseudomonas aeruginosa.
Keywords: Complementation; DNA repair; Homologous recombination; RecA.
Abbreviations: X. c. pv. citri, Xanthomonas campestris pathovar citri; LB, Luria-Bertani; PCR, polymerase chain reaction; UV, ultraviolet; CFU, colony-forming unit; MMS, methylmethane sulfonate; ORF, open reading frame; Kmr, kanamycin-resistance gene; RF, replicative form.
Introduction
Xanthomonas campestris pv. citri, a Gram-negative, non-spore-forming bacterium, and one of approximately 140 pathovars of X. campestris (Verniere et al., 1993), causes citrus canker disease and thereby imposes an enormous economic burden worldwide. To date, only one gene, referred to as pthA, has been associated with the virulence of this organism (Lawson et al., 1989; Swarup et al., 1991, 1992); additional virulence genes remain to be identified. However, molecular studies of X. c. pv. citri have been hampered by the lack of recombination-deficient mutants. Thus, it has been difficult to maintain recombinant plasmids in X. c. pv. citri without the occurrence of deletions or rearrangements in the plasmid, which are likely the result of homologous recombination.
The RecA protein plays a major role in homologous recombination (Camerini-Otero and Hsieh, 1995; Clark, 1973; Cox, 1991; Kowalczykowski et al., 1994; Ogawa and Ogawa, 1986; Roca and Cox, 1990). More than 60 recA genes have been isolated and characterized from a wide variety of organisms, including X. c. pv. oryzae and X. c. pv. campestris (Karlin and Brocchieri, 1996; Lee et al., 1996;
Lloyd and Sharp, 1993; Miller and Kokjohn, 1990; Rabibhadana et al., 1993). We have now cloned, sequenced, and characterized the recA gene of X. c. pv. citri as well as constructed a recA mutant of this organism.
Materials and Methods
Bacterial Strains and Plasmids
The bacterial strains and plasmids used in this study are listed in Table 1. Both X. campestris and E. coli strains were grown in Luria-Bertani (LB) medium at 28°C and 37°C, respectively. Chloramphenicol, kanamycin, and gentamycin were added to LB medium at final concentrations of 30, 50, and 15 µg/ml, respectively.
DNA Manipulation and Hybridization
Plasmid DNA was isolated from bacterial cultures by the method of Birnboim and Doly (1979), and was further purified by ethidium bromide-CsCl density gradient centrifugation. Chromosomal DNA was isolated from X. c. pv. citri as described by Pitcher et al. (1989). Restriction enzyme digestions and ligation were carried out according to standard methods described by Sambrook et al. (1989). The competent X. c. pv. ciri cells were prepared and subjected to electroporation by the method of Yang
1Corresponding author. Tel: +886-2-29031111 ext. 2462; Fax: +886-2-29021124; E-mail: bio1004@mails.fju.edu.tw