Bot. Bull. Acad. Sin. (2000) 41: 143-149
Chi In vitro pollination of lily ovules
Interspecific crosses of lily by in vitro pollinated ovules
H.S. Chi1
Department of Horticulture, National Chiayi University, Chiayi 600, Taiwan, Republic of China
(Received June 4, 1999; Accepted November 25, 1999)
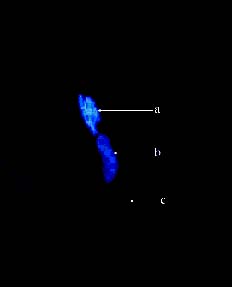
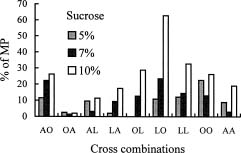
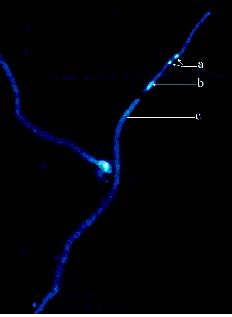
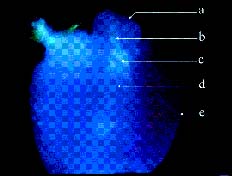
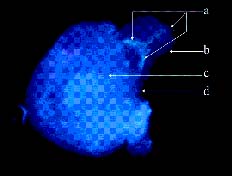
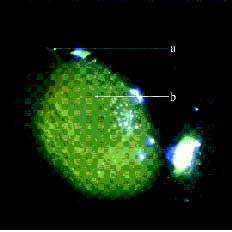
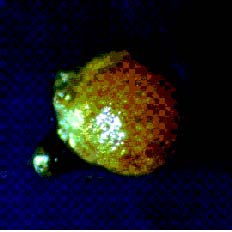
Abstract. Because of pre- and post-fertilization barriers, successful interspecific crosses have not been reported for Lilium using Asiatic hybrid as the mother to cross with Oriental or Lilium longiflorum. Using an in vitro ovular pollination method, where pollen tubes directly penetrate isolated ovules, this study demonstrated that 1) the Brewbaker and Kwack agar plate containing sucrose (10%), with the isolated ovules being placed ca. 1 mm apart from the pollen, is optimal for pollination, 2) interspecific crossing barriers do not occur during penetration of the pollen tubes into the micropyle, 3) micropylar penetration can be obtained in all kinds of intra- and inter-specific cross combinations of lily cultivars, 4) multiple penetration occurs, in which the penetrating pollen tubes pass the embryo sac without entering the synergid cells, 5) a small amount of pollen tube penetrates the synergid cells in the A x O cross, and 6) completion of pollen tube penetration (8-12 h after germination) is much earlier than the division of the generative cell in sperm cells (17 h after germination), which may be the primary cause for the failure in fertilization.
Keywords: In vitro ovular pollination; Interspecific crosses; Lilium; Micropylar penetration; Pollen tube; Sperm cells; Synergid.
Abbreviations: A, Asiatic hybrid; O, Oriental hybrid; L, Lilium longiflorum; MP, micropylar penetration; HAG, hour after germination.
Introduction
Genus Lilium comprises about 85 species that are classified into seven sections, and more than half of these species originate from Asia (Beattie and White, 1993; De Jong, 1974). There are three important modern commercial lilies: the Asiatic hybrid (A), derived from crosses within the Sinomartagon section), the Oriental hybrids (O), derived from crosses within the Archelirion section), and the longiflorum group (L), (Van Creij et al., 1993). Most of the commercial cultivars were bred and selected in The Netherlands (Van Tuyl et al., 1997), and are only suitable for cultivation in a moderate climate. Owing to this, the subtropical lily industry is hampered by the plants poor tolerance of subtropical culture conditions, which causes poor bulb growth and low quality cut flowers (Chin et al., 1996). Therefore, widening the genetic basis by introducing heat-tolerance together with disease resistance (the botrytis resistance of Orientals, the virus resistance of Asiatics, and the fusarium resistance of trumpets) and special flower colors and forms from wild species into commercial lily assortments is very important in subtropical lily breeding programs (Chi et al., 1999). For these purposes, interspecific crosses have to be made. Some of these interspecific crosses in the groups L x A, L x O, O x L, and O x A have been successful (Van Tuyl et al., 1997),
but the A x L and A x O crosses have not. Therefore, no hybrid plant of the latter crossing types has been obtained, even though different in vitro methods have been exploited.
It has been proposed that the limiting hindrance factors of interspecific crosses in Lilium are primarily fertilization barriers that can be separated into pre-and post-fertilization barriers (Van Tuyl et al., 1991). To overcome pre-fertilization barriers, the cut-style method, the grafted style method, the placenta pollination method, and the in vitro ovule pollination method have been tested (Asano and Myodo, 1977a, b; Janson et al., 1993; Van Tuyl et al., 1991; Willemse et al., 1995). Embryo rescue, ovary-slicing, and ovule culture have been tried to overcome post-fertilization barriers (Asano, 1980; Okazaki et al., 1992, 1994; Van Tuyl et al., 1991). By overcoming both barriers, interspecific lily crosses have been made more efficient, resulting in increased numbers of hybrid plantlets in a single interspecific cross and more successful interspecific combinations (Van Tuyl et al., 1991, 1997).
Recently, in an L x A cross using the in vitro ovular pollination method, where L was L. longiflorum Gelria and A was the Asiatic hybrid Enchantment, Willemse et al. (1995) showed micropylar penetrations of 8-17%; however, fertilization was not successful (Plyushch et al., 1995; Willemse et al., 1995). The purpose of this research was to optimize the conditions for in vitro ovular pollination and to study at which stage the barriers may occur in those crosses.
1Corresponding author. Tel/Fax: +886-5-2777484; E-mail: hschi@mail.ncyu.edu.tw