Bot. Bull. Acad. Sin. (2000) 41: 175-182
Lee et al. A soybean seed maturation gene
Promoter activity of a soybean gene encoding a seed maturation protein, GmPM9
Pei-Fang Lee1, Yue-Ie C. Hsing2 and Teh-Yuan Chow1, 2, 3
1Graduate Institute of Botany, National Taiwan University, Taipei, Taiwan, Republic of China
2Institute of Botany, Academia Sinica, Nankang, Taipei, Taiwan, Republic of China
(Received November 18, 1999; Accepted January 17, 2000)
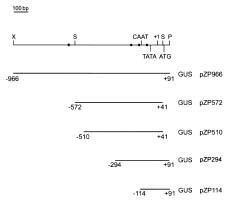
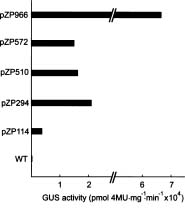
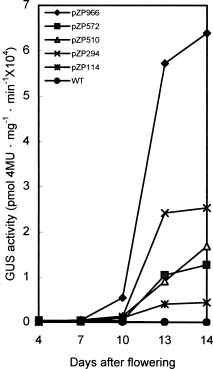
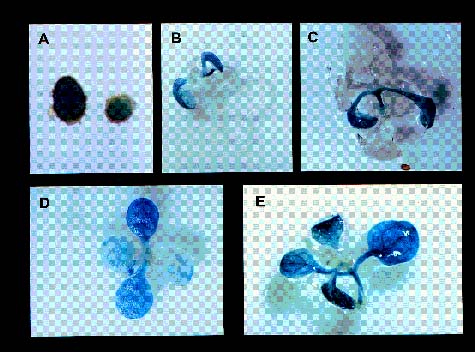
Abstract. Late embryogenesis abundant (LEA) proteins are synthesized during the late stages of seed development and have been widely reported in monocot and dicot plants. In order to understand the developmental regulation of the promoter of a soybean gene encoding LEA protein GmPM9, we fused a series of fragments of the promoter region to the b-glucuronidase (GUS) reporter gene (uirA) and transformed the resulting constructs (pZP966, pZP572, pZP510, pZP294 and pZP114) into Arabidopsis thaliana. GUS enzyme activities were detected in mature seeds and cotyledons and hypocotyls of seedlings in transgenic plants containing any one of the five constructs; they were not detected in other tissues at different developmental stages. During seed development, GUS began to appear at 10 days after flowering (DAF) and increased rapidly to a maximum in the mature seeds at 14 DAF. The longest promoter construct (pZP966) enabled the transgenic plants to exhibit the highest GUS activity, whereas the shortest construct (pZP114) was sufficient to direct the expression of the GUS gene at a detectable level. The above findings indicate that the promoter of gGmPM9 can be used to express desirable genes in seeds during late seed maturation. The expression of the GUS gene could be induced in the leaves of the transgenic plants containing pZP966 by salt and desiccation, but not by ABA treatment.
Keywords: b-glucuronidase; Late embryogenesis abundant; Promoter; Transgenic Arabidopsis.
Abbreviations: ABA, abscisic acid; DAF, days after flowering; GUS, b-glucuronidase; LEA, late embryogenesis abundant; MU, methylumbelliferone; MUG, methylumbelliferyl glucuronide; PEG, polyethylene glycol.
Introduction
Late embryogenesis abundant (LEA) genes are highly expressed at the late stage during seed development. They have been studied extensively and isolated from many plants, including monocots and dicots (Baker et al., 1988; Close et al., 1989; Gaubier et al., 1993; Hsing et al., 1995; Mundy and Chua, 1988; Pla et al., 1991). LEA proteins have been classified into at least six groups based on their conserved sequences (Dure, 1993). In addition to being expressed in the embryo, some LEA genes can be induced in vegetative tissues by osmotic stress, including salt treatment and desiccation, and exogenous abscisic acid (Chandler and Robertson, 1994; Skriver and Mundy, 1990). LEA proteins are believed to play a role in cellular protection because they share the extremely hydrophilic property as well as the timing for accumulation. For example, expression of the group 3 LEA protein, HVA1, from barley, conferred a tolerance to water deficit and salt stress in transgenic rice (Xu et al., 1996).
In addition to determining the function of LEA proteins, it is important to investigate the molecular mechanisms that regulate LEA genes. A number of LEA genes are thought
to respond to ABA, and their promoters contain conserved motifs called ABREs (ABA-responsive elements), which interact with nuclear protein factors. Guiltinan et al. (1990) reported that EmBP-1, a leucine-zipper DNA binding protein, could bind to the ABRE sequence (CACGTGGC) of the wheat Em gene. In order to analyze the regulation mechanism of LEA genes, several promoters from different plants were fused to b-glucuronidase (GUS) reporter gene (uirA) (for instance, Goupil et al., 1992; Hull et al., 1996; Michel et al., 1994; Rouse et al., 1996; Yamaguchi-Shinozaki and Shinozaki, 1993). These studies indicated that the promoters of different LEA genes would respond to one or more stimuli, including ABA, cold, salt, or desiccation. These promoters could direct GUS expression to mature seeds, pollens, roots, stems or trichomes, with most of these LEA genes expressing in seeds. Promoter deletion analysis revealed a variety of cis-acting sequences within these promoters that conferred differential responsiveness to environmental stimuli (Giraudat et al., 1994; Hull et al., 1996).
We isolated and characterized gGmPM9 (GmPM, Glycine max physiological mature) (Lee et al., 1992), a genomic sequence from soybean. This sequence contained: a 1 kb, 5' untranscripted sequence, a 0.3 kb, 3' untranslated sequence, and coded for the 16 kDa seed maturation protein, GmPM9, which was highly homologous to
3Corresponding author. Tel: 886-2-2789-9590 ext. 220; Fax: 886-2-2782-7954; E-mail: tychow@gate.sinica.edu.tw