Cheng and DaiSequence and transcript analysis of gene cluster orf140-nad3-rps12-atp1 in mung bean seedling mitochondria
Bot. Bull. Acad. Sin. (2000) 41: 183-190
Sequence and transcript analysis of the orf140-nad3-rps12-atp1 gene cluster in the mitochondrial DNA of mung bean (Vigna radiata L. (Wilzek) cv. Tainan No. 5)
Yuchang Cheng and Hwa Dai1
Institute of Botany, Academia Sinica, Nankang, Taipei, Taiwan 115, ROC
(Received July 26, 1999; Accepted January 26, 2000)
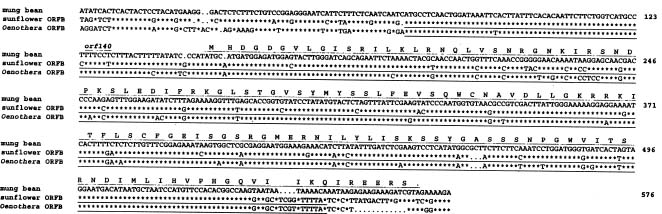
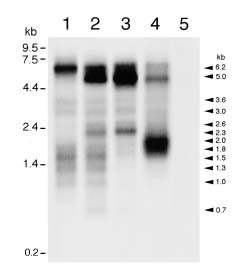
Abstract. An orf140-nad3-rps12-atp1 gene cluster, which spans a 7.1 kb region of the mung bean mitochondrial genome, was identified and completely sequenced. The coding regions of nad3, rps12, and atp1 genes are highly conserved between mung bean and other plant species. The nucleotide sequences of orf140 and its 5'-flanking region are homologous with ORFBs in sunflower and Oenothera, and are also found to be conserved in several chimeric ORFs of various plant CMS lines. The region 3'-downstream from the mung bean atp1 is similar to its counterparts in soybean and common bean, which has been shown to be specific to a subset of legumes. Genes of this cluster are present as a single copy in mung bean. Northern-blot hybridization and RT-PCR suggested that the four genes are co-transcribed and then processed into transcripts of different sizes. Variation of accumulation levels of different transcripts was observed.
Keywords: DNA sequence; Mitochondrial gene cluster; Mung bean; orf140-nad3-rps12-atp1; Transcription.
Introduction
The higher plant mitochondrial (mt) genomes, ranging in size from 208 to 2,400 kb, are much larger and more complicated than their counterparts in animals and fungi (Palmer and Herbon, 1987; Ward et al., 1981). In spite of variation in genome sizes, most plant mt genomes encode roughly the same complement of gene products, including 3 rRNAs, 15-20 tRNAs, and approximately 20 polypeptides (Wolstenholme and Fauron, 1995). Although coding regions of individual genes are highly conserved, the mt genomes of higher plants exhibit distinctive gene orders in virtually every species so far examined (Palmer, 1992). The rearrangements are mainly due to recombination events between repeats of various sizes, which are dispersed throughout the plant mitochondrial genome (Small et al., 1989). Besides the effect on genome organization, frequent genomic recombinations can also create chimeric or truncated open reading frames (ORFs), affect transcriptional patterns, and place genes that are far apart in one genome into close proximity in another species (Hanson and Folkerts, 1992).
Most plant mitochondrial genes are monocistronic and separated by long spacers (Hanson and Folkerts, 1992). When transcripts are analyzed, the mRNAs are found to be generally larger than the actual coding regions and to contain extended non-coding 5'- and 3'-transcribed regions (Binder et al., 1996). Thus, genes located closely together
are inevitably co-transcribed. Examples of mitochondrial gene co-transcription have been reported in a number of plants, including maize (Gualberto et al., 1991), wheat (Gualberto et al., 1988), rice (Nakazono et al., 1995), Arabidopsis (Brandt et al., 1992), petunia (Pruitt and Hanson, 1989) and tobacco (Bland et al., 1986). Most arrays of co-transcribed gene clusters are specific to the mitochondria of a given plant species, except the three gene clusters rrn18-rrn5, nad3-rps12, and rps3-rpl12, which have been highly conserved (Palmer, 1992). The co-transcription of genes that encode proteins functioning in different pathways indicates that post-transcriptional and post-translational regulation are involved in controlling the abundance of the gene products. Furthermore, the high variability of genome organizations between different species and the complexity of possible transcription patterns make extensive sequencing and transcription mapping necessary to understand the mechanisms involved in regulation of plant mitochondrial gene expression.
In this work, we have completely sequenced a 7.1 kb region of mung bean mtDNA, which contains an unidentified ORF (orf140), nad3, rps12 and atp1 genes. We have also shown that these four genes are co-transcribed.
Materials and Methods
Plant Materials
Mung bean (Vigna radiata L. (Wilzek) cv. Tainan No. 5) was used as the source for mtDNA and mtRNA purification. Etiolated seedlings were grown in darkness at 30°C on vermiculite soaked in water.
1Corresponding author. Tel: (02) 2789-9590-326; Fax: (02) 2782-7954; E-mail: bodaihwa@gate.sinica.edu.tw