Bot. Bull. Acad. Sin. (2000) 41: 191-196
Hou et al. Dioscorins with both carbonic anhydrase and trypsin inhibitor activities
Dioscorins from different Dioscorea species all exhibit both carbonic anhydrase and trypsin inhibitor activities
Wen-Chi Hou, Hsien-Jung Chen and Yaw-Huei Lin1
Institute of Botany, Academia Sinica, Nankang, Taipei 115, Taiwan, Republic of China
(Received January 31, 2000; Accepted February 24, 2000)
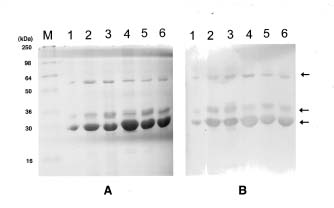
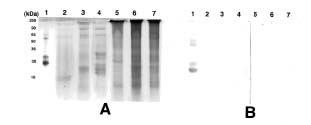
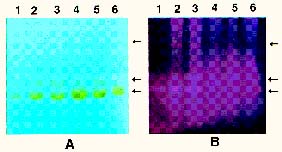
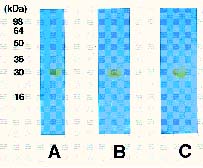
Abstract. Dioscorins, the storage proteins of yam tubers, from six cultivars of three Dioscorea species including D. batatas, D. alata, and D. pseudojaponica were detected in crude extracts of yam tubers but not leaves by immuno stainings on PVDF membrane with a polyclonal antibody which was raised against 28 kDa dioscorin from Dioscorea batatas Decne. It was demonstrated that the dioscorins from the above-mentioned yam species exhibited both carbonic anhydrase (CA) and trypsin inhibitor activities by activity stainings on 15% SDS-PAGE gels. Dioscorins were identified as glycoproteins by ConA-peroxidase activity staining. Dioscorins from D. batatas Decne contained no zinc atoms detectable by atomic absorption spectrophotometry. Neither 5 mM acetazolamide nor 2,6-pyridinedicarboxylic acid inhibited the CA activity of dioscorins.
Keywords: Acetazolamide; Carbonic anhydrase; Dioscorin; Pyridinedicarboxylic acid; Storage proteins; Trypsin inhibitor; Yam.
Abbreviations: CA, carbonic anhydrase; 2-ME, 2-mercaptoethanol; PVDF, polyvinylidene difluoride; SDS-PAGE, sodium dodecylsulfate-polyacrylamide gel electrophoresis; TI, trypsin inhibitor.
Introduction
Carbonic anhydrases (CAs) catalyze the simple interconversion of CO2 and HCO3-. They are found in almost all organisms and are notable for extremely high turnover numbers (Tashian, 1989; Badger and Price, 1994). In cyanobacteria and plants, CAs facilitate the interchange of CO2 and HCO3- and play a key role in CO2 fixation of photosynthesis (Hatch and Burnell, 1990; Badger and Price, 1992). In mammals, CAs also facilitate the interchange of CO2 and HCO3- and play a key role in respiration (Tashian and Hewett-Emmett, 1984; Forster, 1988). According to Hewett-Emmett and Tashian (1996), CA is encoded by three independent CA gene families (a-CA, b-CA, and g-CA). Most CAs of mammals and green algae belonged to the a-CA family, those of plants and eubacteria to the b-CA family, and those of archaebacteria to the g-CA family. Surprisingly, based on the cDNA deduced amino acid sequence alignment, the storage protein of yam (Dioscorea cayenensis L.) tuber, dioscorin, was found to be a a-CA related protein (Hewett-Emmett and Tashian, 1996). However, Hewett-Emmett and Tashian (1996) also mentioned that dioscorin was unlikely to have classical CA activity due to the active site alternations.
Patatin, the potato tuber storage protein, has been reported to contain both lipid acyl hydrolase and acyltransferase activities, both of which are involved in tuber tissue response to wounding (Andrews et al., 1988). The soybean vegetative storage proteins VSP a and VSP b both have acid phosphatase activity (Dewald et al., 1992). We have demonstrated that the root storage proteins of sweet potato not only have trypsin inhibitor (TI) activity (Hou and Lin, 1997a), but also dehydroascorbate reductase and monodehydroascorbate reductase activities, which might be involved in response to environmental stresses (Hou and Lin, 1997b). Tarin, the storage protein of taro (Colocasia esculenta L. Schott), was homologous to mannose-binding lectin and curculin in deduced amino acid sequences (Bezerra et al., 1995). Recently, it was reported that yam bean (Pachyrhizus erosus L. Urban) storage proteins YGB 1 and YGB 2 exhibited cysteine protease activities and YBP 2 showed sequence homology to protease inhibitors (Gomes et al., 1997). We reported recently that the purified dioscorins from tubers of Dioscorea batatas Decne (imported from Japan) showed both CA and TI activities (Hou et al., 1999a) and both dehydroascorbate reductase and monodehydroascorbate reductase activities (Hou et al., 1999b). In this report, we extend the work to show that both CA and TI activities could be found in dioscorin samples from six cultivars of three Dioscorea species including D. batatas, D. alata and D. pseudojaponica grown in Taiwan by activity staining on 15% SDS-PAGE gels. Some properties of dioscorins were also included.
1Corresponding author. Tel: 886-2-2789-9590 ext. 320; Fax: 886-2-2782-7954; E-mail: boyhlin@ccvax.sinica.edu.tw