Bot. Bull. Acad. Sin. (2000) 41: 203-212
Lai et al. Papaya axillary shoots enhanced by ethylene
Enhancement of papaya axillary shoot proliferation in vitro by controlling the available ethylene
Chuo-Chun Lai1, Shyi-Dong Yeh2, and Jiu-Sherng Yang1,3
1Department of Botany, National Chung-Hsing University, Taichung 402, Taiwan, Republic of China
2Department of Plant Pathology, National Chung-Hsing University, Taichung 402, Taiwan, Republic of China
(Received June 22, 1999; Accepted December 31, 1999)
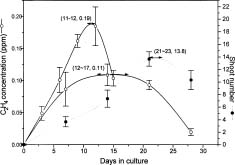
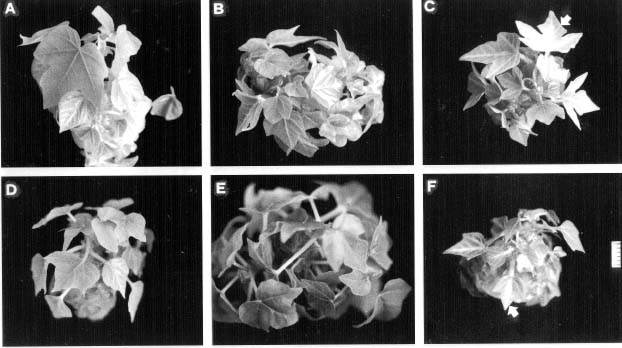
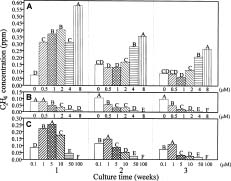
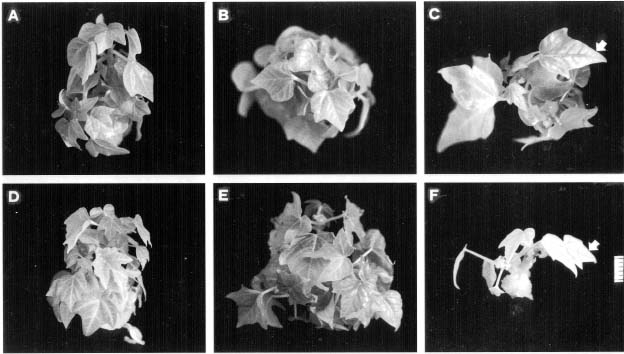
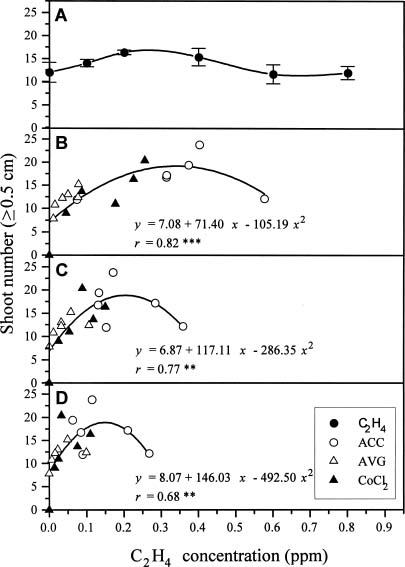
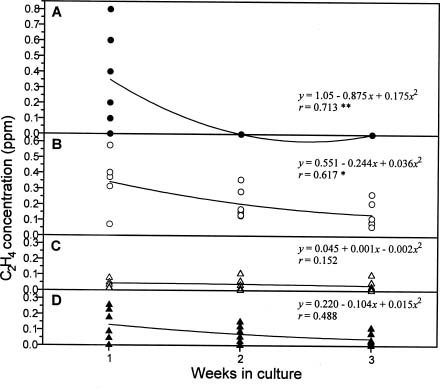
Abstract. The timing of ethylene concentration favorable for in vitro proliferation of papaya (Carica papaya L.) multiple shoot clusters through outgrowth of axillary buds was investigated. Exogenous ethylene in culture flasks was regulated during the first week by using a gas diffusion equilibration procedure, which used a transparent box as an indirect space for the ethylene injection and flasks sealed with 0.02 µm filters for diffusion and equilibration. There was a 36% increase in shoot number and a 50% increase of leaf number by adding 0.2 or 0.4 ppm ethylene into the box the first week, followed by aerating the flasks the following two weeks. The level of endogenous ethylene was also modified by adding the ethylene biosynthesis precursor (ACC) and the inhibitors (AVG and CoCl2) to the culture medium in the flasks under sealed conditions. The greatest enhancement rate of shoot number (75%) was achieved with 2 µM ACC. Shoot proliferation rates by applying 0.5 µM AVG and 5 µM CoCl2 were enhanced by 23% and 49%, respectively. The relationships between shoot number and ethylene concentration during a three-week incubation period were analyzed to reveal the most favorable ethylene concentrations. Concentrations with the best proliferation were 0.34, 0.20 and 0.15 ppm for Weeks 1, 2 and 3, respectively. The results indicated that a relatively higher ethylene level during the early incubation period, followed by low levels subsequently might be most favorable for improved rates of papaya axillary shoot proliferation.
Keywords: ACC; AVG; Carica papaya; Cobalt chloride; Ethylene; Gas diffusion equilibration procedure.
Abbreviations: ACC, 1-Aminocyclopropane-1-carboxylic acid; AVG, Aminoethoxyvinylglycine; BAP, 6-Benzylaminopurine; MACC, 1-(Malonylamino) cyclopropane; NAA, a-Naphthaleneacetic acid; PP, Polypropylene.
Introduction
Papaya (Carica papaya L.) is an economically important fruit crop of the tropics and subtropics that is highly susceptible to the papaya ringspot virus (PRSV). Currently, the most promising method for overcoming plant viral diseases is genetic engineering. Production of PRSV resistant papaya plants via introduction of the PRSV coat protein gene has been reported (Cheng et al., 1996). What is needed now is a rapid clonal propagation protocol to increase the virus-resistant transgenic genotypes for commercial planting. Plant multiplication in tissue culture by axillary bud proliferation has proved suitable for papaya clonal multiplication (Litz and Conover, 1978). The earlier papaya micropropagation studies established the details with respect to sexual type selection, the season for explanting (Litz and Conover, 1981), source of explants, and hormone components of culture media (Litz and Conover, 1978; Drew, 1988). Nevertheless, the shoot proliferation rates have remained generally low.
Plant growth and development in vitro is governed not only by the composition of the culture medium, but also by constituents of the culture vessel atmosphere, such as carbon dioxide, oxygen, and especially, the plant growth regulator, ethylene (Blazková et al., 1989; Buddendorf-Joosten and Woltering, 1994). The effects of ethylene on in vitro culture have been studied in relation to flowering (Blazková et al., 1989), rooting (Pérez-Bermúdez et al., 1985), and axillary shoot development (Nour and Thorpe, 1994). The ethylene levels were controlled by either supplementing the gas exogenously (Kevers et al., 1992) or by experimenting with various types of culture vessels and closures to control the accumulation of endogenously evolving ethylene (Blazková et al., 1989). Dimasi-Theriou and Economou (1995) found that the application of 0.1 ppm ethylene during the first two weeks of culture increased the number and length of peach axillary shoots, but that excessive ethylene supplements inhibited shoot growth. A similar finding was reported by Lai et al. (1998) for axillary plant proliferation of papaya. Compared to continuously unaerated cultures, a 41% increase in the number of new shoots was obtained when endogenous ethylene was permitted to accumulate in culture vessels to 0.06 ppm during the first week of culture, followed by aerating the vessels the subsequent two weeks.
3Corresponding author. Tel: 886-4-2840417-404; Fax: 886-4-2874740; E-mail: jsyang@dragon.nchu.edu.tw