Bot. Bull. Acad. Sin. (2000) 41: 213-218
Kao et al. C-13 of a subalpine forest
Vertical profiles of CO2 concentration and d13C values in a subalpine forest of Taiwan
Wen-Yuan Kao1, Yu-Shiun Chiu, and Wan-Hwa Chen
Institute of Botany, Academia Sinica, Taipei, Taiwan, Republic of China
(Received September 20, 1999; Accepted January 28, 2000)
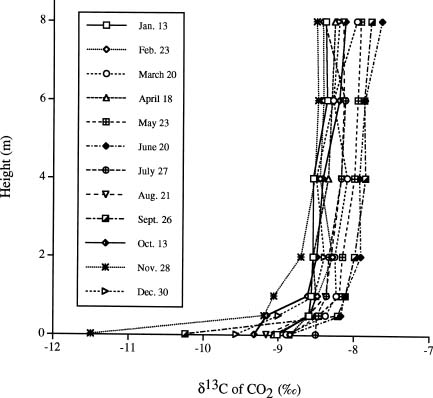
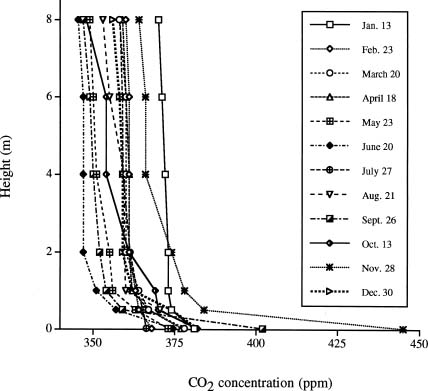
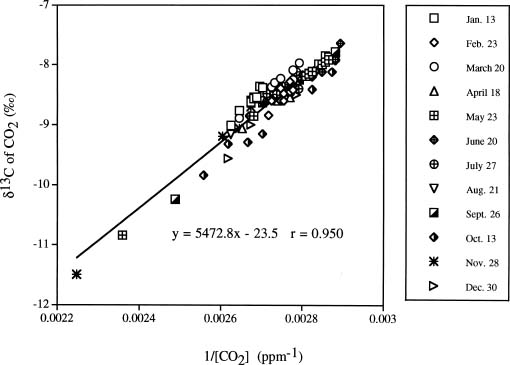
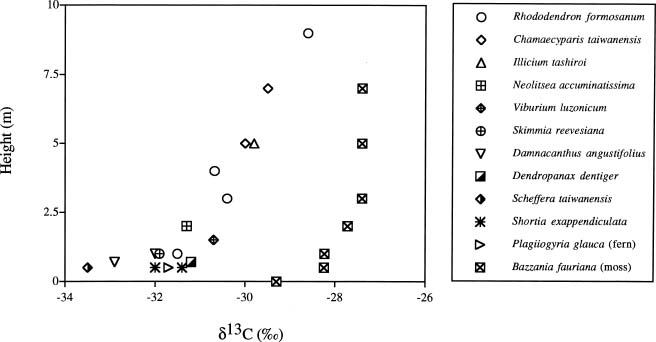
Abstract. We investigated the vertical gradients in CO2 concentration and stable carbon isotope ratio (d13Cco2) of the canopy air within a coniferous-hardwood, Chamaecyparis and Rhododendron dominated, subalpine forest in Taiwan. The stable carbon isotope ratio of vascular plants and a epiphytic bryophyte species (Bazzania fauriana) (d13Cleaf) from different heights within the forest were also analyzed. Results revealed that CO2 and d13Cco2 gradients did exist within the forest, with higher CO2 concentrations and more negative d13Cco2 values in air sampled from the lower canopy. The average vertical gradients in CO2 and d13Cco2 value of the CO2 of the 12 sampling dates were 28.5 ± 6.1 ppm and 1.3 ± 0.3, respectively. Seasonal patterns of the relationship between 1/[CO2] and the corresponding d13C were pronounced, with steady decreases in the slopes and increases in the intercepts found from January to August. A decreasing d13Cleaf with decreasing height was also measured in a bryophyte (ranging from -27.4 to -29.2), canopy and understory leaves (ranging from -28.6 to -33.5). It was estimated that photosynthetic physiology affected by microclimates within the forest contributed approximately 2.8 of variation of the vertical gradient of vascular plant d13C values.
Keywords: Bryophyte; CO2 ; Subalpine forest; Stable carbon isotope ratio.
Introduction
Forest ecosystems are an important carbon pool and have profound impacts on atmospheric CO2 concentrations. In particular, soil-respired CO2 in forests has been reported as a significant component in global carbon cycling (Woodwell et al., 1983). The CO2 released during respiration and decomposition may diffuse through the forest canopy into the atmosphere or a fraction of this CO2 may be reassimilated through photosynthesis by the forest ecosystem (Lloyd et al., 1996; Sternberg et al., 1989). Thus, there are two major sources of CO2 for photosynthesis within forests: one is from bulk air and the other from soil respiration.
Internal carbon fluxes within forest canopies and their interactions with soil and atmospheric exchange processes can be addressed using carbon isotopes. The mean value of atmospheric CO2 is currently -8 but varies seasonally in response to the patterns of photosynthesis and respiration (Conway et al., 1994; Mook et al., 1983). Photosynthesis discriminates against 13CO2, thus plants have a lighter carbon isotopic composition in their tissue in comparison to the atmospheric CO2. The respired CO2 derived from root respiration and decomposition of soil organic matter has a d13C value close to that of the organic matter of the dominant species in the forest community
(Flanagan et al., 1996). Accordingly, the two sources of CO2 within the forest canopy have different isotopic signals.
Vertical gradients in CO2 and d13C values have been studied in different forest ecosystems in different areas of the world. Turbulent mixing between the two sources of CO2 within the canopy and discrimination against 13CO2 during photosynthesis results in d13C of ratios of canopy air that are more depleted near the soil surface than at top of the canopy (Broadmeadow et al., 1992; Buchmann et al., 1997a,b; Flanagan et al., 1996; Francey et al., 1985; Quay et al., 1989; Sternberg, 1989; Van der Merwe and Medina, 1989). The range of the gradients depends on forest development, forest structure, and forest types (Buchmann et al., 1997b). To our knowledge, no similar study has been done in any forests of Taiwan. Since 1992, a long-term ecological study (LTER) has been set-up at a subalpine ecosystem within a natural preserve. The LTER study emphasizes the structure and function of the forest ecosystem as well as its carbon and nutrient flux. Understanding variations in the concentration and isotopic composition of CO2 within and above vegetation could provide insights into ecosystem functioning (Lloyd et al., 1996). In the present study, we investigated the profiles of CO2 concentration and d13C values of the canopy air and vegetation at different heights of the forest stand to understand processes related to carbon flux within the ecosystem. Stable carbon isotope ratios of vascular plants and a moss species (d13Cleaf) from different heights within the forest were also analyzed. The objectives of the present study were to understand: (1) whether a vertical profile in CO2
1Corresponding author. Phone: 886-2-27899590; Fax: 886-2-27827954; E-mail: bowykao@ccvax.sinica.edu.tw