Bot. Bull. Acad. Sin. (2000) 41: 225-230
Gu and Ko Parasexual cycle in Phytophthora parasitica
Occurrence of parasexual cycle in Phytophthora parasitica
following protoplast fusion
Y.H. Gu1 and W.H. Ko2
Department of Plant Pathology, Beaumont Agricultural Research Center, University of Hawaii at Manoa, Hilo, Hawaii 96720, USA
(Received August 12, 1999; Accepted February 14, 2000)
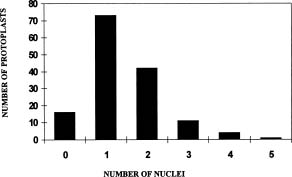
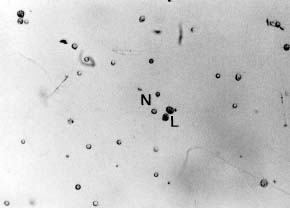
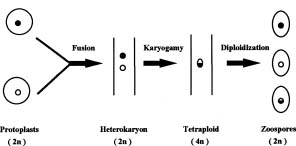
Abstract. Metalaxyl-resistant (Mr) and chloroneb-resistant (Cnr) protoplasts of Phytophthora parasitica were fused, with heterokaryon formation rates ranging from 2.6 × 10-3 to 8.2 × 10-3. The heterokaryosis was confirmed in six of seven putative heterokaryons by the production of both Mr zoospores and Cnr zoospores, and/or Mr + Cnr zoospores. The 7th putative heterokaryon produced only Cnr zoospores, probably because of the loss of the Mr chromosome during subculture. The heterokaryons produced normal-sized zoospores as well as large zoospores that were almost twice the size of normal-sized zoospores. Fifty percent of the normal-sized zoospores contained Mr and Cnr genes. The results suggested an occurrence of diploidization after karyogamy of the heterokaryon and thus demonstrated the completion of events leading to a parasexual cycle following protoplast fusion.
Keywords: Diploidization; Phytophthora parasitica; Parasexual cycle; Protoplast fusion.
Introduction
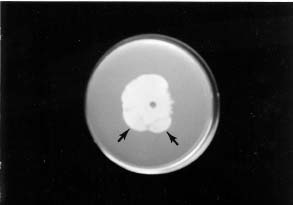
Protoplast fusion has proved to be a highly efficient procedure for obtaining heterokaryons and enabling the parasexual cycle of filamentous fungi readily available for genetic studies and for industrial strain improvement (Anne and Peberdy, 1985; Peberdy, 1989). The procedure is especially valuable for Phytophthora because of the rare occurrence of hyphal fusion in this group of fungi (Judelson and Yang, 1998). However, the number of reports on this subject is very limited for Phytophthora (Layton and Kuhn, 1988; Lucas et al., 1990). Protoplast fusion has been used by Lucas et al. (1990) to generate somatic hybrids of Phytophthora capsici Leonian. Double drug-resistant strains were obtained when protoplasts of a metalaxyl resistant mutant of Phytophthora sojai Kaufmann & Gerdemann were fused with fluorotryptophan-resistant protoplasts (Layton and Kuhn, 1988). Fifteen of the hybrids produced zoospores of each parental phenotype, indicating a heterokaryon formation, while eleven hybrids produced a large number of zoospores that were resistant to both metalaxyl and fluorotryptophan, suggesting that karyogamy had possibly occurred. However, evidence that the selective markers represented nuclear genes has been lacking (Layton and Kuhn, 1988).
Chang and Ko (1990) obtained mutants of Phytophthora parasitica Dastur that were resistant to either metalaxyl or chloroneb and found that metalaxyl resistance and chloroneb resistance in these mutants were each conferred by a single dominant gene in heterozygous condition. Because of the availability of these nuclear gene markers, we used the mutants to perform protoplast fusion. Afterwards, we were able to detect events leading to a parasexual cycle, similar to those occurring in haploid fungi (Burnett, 1975; Pontecorvo, 1956). Details of the study are reported here.
Materials and Methods
Mutant Selection
Single-zoospore isolates P991 and 6134 of P. parasitica (Chang and Ko, 1990) were cultured on V-8 agar (10% V-8 juice, 0.02% CaCO3 and 2% Bacto agar) containing either 25 ppm metalaxyl (subdue 2E, 25.1% active) or 200 ppm chloroneb (Terraneb WP, 65% active). Fast-growing sectors that appeared after 3 weeks were transferred to the same fungicide medium. Single-zoospore cultures were isolated from those that retained resistance after five successive subcultures, and zoospore cultures that displayed resistance after four subcultures on fungicide-free medium were selected. Fungicide resistance was determined by placing culture discs (4 mm) on V-8 agar amended with 25 ppm metalaxyl or 200 ppm chloroneb. Cultures that were sensitive to metalaxyl did not grow in 7 days at 24°C on medium containing metalaxyl, while those that were resistant grew continuously, up to 18-20 mm in diameter, during the same incubation period. Similarly, cultures that
1Present address: USDA, ARS, Tree Fruit Research Laboratory, Wenatchee, Washington 98801, USA
2Corresponding author. Tel: (808) 974-4105; Fax: (808) 974-4110.