Bot. Bull. Acad. Sin. (2000) 41: 257-262
Huang et al. RAPD markers of Chrysanthemum hybrids
Genetic analysis of Chrysanthemum hybrids based on RAPD molecular markers
Sheng Chung Huang1, Chi Chu Tsai, and Chian Shinn Sheu
Taichung District Agricultural Improvement Station, Changhua, Taiwan, Republic of China
(Received August 23, 1999; Accepted May 10, 2000)
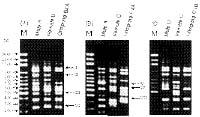
Abstract. Forty-five random primers were screened, of which twenty-two primers were selected to detect the molecular marker in three hybrid combinations of Chrysanthemum by using random amplified polymorphic DNA (RAPD). From this study, the patterns of molecular markers could be classified into seven types: Type I markers shared bands in both parents, and offspring; Type II markers shared bands in male and female parents; Type III markers shared bands in male parent and offspring; Type IV markers shared bands in female parent and offspring; Type V markers were presented in the male parent only; Type VI markers were present in the female parent only; Type VII markers were present in offspring only. Of these, only Type III markers were suitable for identifying the true male parent. Different unique markers of Type VII in offspring are quite suitable as identifying markers of new hybrids to protect the rights of plant breeders. In this study, 34.4% to 48.9% of the RAPD markers were found to reveal additivity among parents and offspring in three hybrid combinations of Chrysanthemum. However, 38% to 52.6% of markers (Type II, V, and VI) were absent in offspring, but 11.6% to 13.1% of unique markers (Type VII) were present in offspring. Moreover, there were no definite rules as to whether markers in offspring were more similar to female or to male parents by similarity analysis. In two hybrid combinations, the parents were more similar to each other than either was to the offspring. The above results illustrate that the genetics of Chrysanthemum are very complex. RAPDs, however, are a powerful tool to detect different molecular markers in hybrid populations of Chrysanthemum cultivars.
Keywords: Chrysanthemum; Hybrids; Molecular marker; RAPD.
Introduction
Chrysanthemum morifolium Ramat (Asteraceae) has been bred for 3,000 years in China and Japan. It is one of the major horticultural crops in the Netherlands today (Wolff et al., 1994). Chrysanthemum morifolium cultivars are polyploids belonging to a hexaploid species with an average chromosome number of 54 (Dowrick, 1953; Langton, 1989), but the exact origin of the hexaploid species is still unknown (Wolff et al., 1994). The species has a strong self-incompatibility system, as do all members of the Asteraceae family (Richards, 1986). It is known that the self-incompatibility in the Asteraceae is determined by a multiallelic sporophytic system. This system is correlated with dry papillate stigmas, trinucleate pollen, and the incompatibility reaction at the stigmatic surface (Richards, 1986), but the genetics of Chrysanthemum have not yet been completely revealed (Wolff and Peters-Van Rijn, 1993; Zagorski et al., 1983). Selfing is generally not possible, although some pseudo-self-incompatible plants have been discovered (Anderson et al., 1992). The rate of successful crosses between related and unrelated cultivars is low, usually only 5% to 50% (Zagorski et al., 1983). Nevertheless, breeding of
Chrysanthemum cultivars has been accomplished by traditional techniques.
However, not all types of markers are suitable for breeding applications. Morphological and cytological markers are not useful for breeding analysis (Roxas et al., 1993). Although isozyme markers are useful to characterize genetic diversity (Fiebich and Henning, 1992; Roxas et al., 1993), and to identify the hybrids of cultivars (Roxas et al., 1993), the paucity of isozyme loci restricts their usefulness in breeding (Helentjaris et al., 1986). DNA markers have been used to manipulate marker-assisted selection (MAS), and to guide the introgression of target genes from related species by restriction fragment length polymorphism (RFLP) in the past several years (Wolff et al., 1994). However, RFLP is labor intensive and costly.
An alternative technique for identifying molecular marks called random amplified polymorphic DNA (RAPD) has been developed (Williams et al., 1990). In this method, by using a single arbitrary primer (10 mer) and amplifying DNA by polymerase chain reaction (PCR), the resulting DNA markers easily can be separated on an agarose gel by electrophoresis (Williams et al., 1990). The advantages of RAPD is its simplicity, rapidity, the requirement for only a small quantity of DNA, and the ability to generate numerous polymorphisms (Cheng et al., 1997). Therefore, it has been a powerful technique for genetic analysis (Chapco et al., 1992; Kiss et al., 1993; Landry et al., 1993; Wight et al., 1993; Williams et al., 1990).
1Corresponding author. Tel: +886-4-8523101 ext. 200; Fax: +886-4-8525841.