Bot. Bull. Acad. Sin. (2000) 41: 263-266
Huang et al. Phase change and ethylene evolution
Ethylene evolution by juvenile and adult developmental phases of Sequoia sempervirens shoots cultured in vitro
Li-Chun Huang1, Ching-I Kuo, Chiu-Hui Wang, Toshio Murashige, and Tan-Chi Huang
Institute of Botany, Academia Sinica, Nankang, Taipei, Taiwan
(Received January 29, 2000; Accepted March 21, 2000)
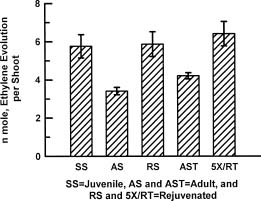
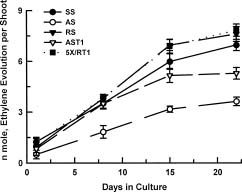
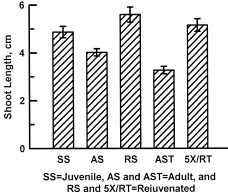
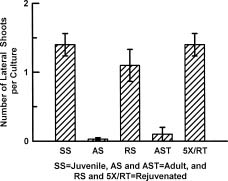
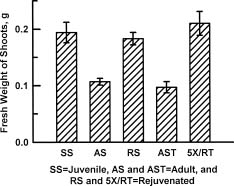
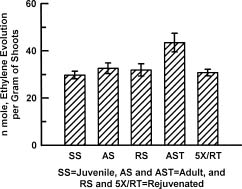
Abstract. Cultures of juvenile and rejuvenated Sequoia sempervirens shoots generated more ethylene than those of adult shoots. But the higher phytohormone production was only indirectly related to the developmental phase. The juvenile and rejuvenated shoots also grew more rapidly; thus, when measured on a per gram tissue basis the rates of ethylene evolution were the same for tissues of both developmental phases, and even higher for one of the adults. The investigation did not establish whether the faster growth of juvenile and rejuvenated shoots was caused by the ethylene; on the other hand, there was no evidence of inhibitory effects.
Keywords: Ethylene; Gas chromatograph; Phase change; Sequoia sempervirens; Tissue culture.
Introduction
Several biochemical markers can now be employed to distinguish the juvenile from the mature or adult developmental phases of certain plants, e.g., differences in peroxidase and esterase isozymes (Brand and Lineberger, 1992; Huang et al., 1996) and in protein phosphorylation (Kuo et al., 1995). Furthermore, the phytohormones gibberellin (Frydman and Wareing, 1973; Rogler and Hacket, 1975a), abscisic acid (Rogler and Hackett, 1975b) and cytokinin (Bouriquet et al., 1985; Mullins et al., 1979) have been shown to be intimately involved in phase change. Gibberellin and cytokinin treatments can cause rejuvenation, whereas abscisic acid can stabilize the adult phase. This investigation was undertaken to determine whether the ubiquitous phytohormone ethylene might also have a role in phase change, inasmuch as it has been associated with diverse growth processes (Buddendorf-Joosten and Woltering, 1994; Dimasi-Theriou and Economou, 1995; Magdalita et al., 1997; Nour and Thorpe, 1994). We measured its production by aseptic cultures of juvenile, adult and rejuvenated shoots of the coastal red-wood tree, Sequoia sempervirens.
Materials and Methods
Shoots for the investigation were obtained from stock cultures established in vitro. Stocks of juvenile shoots were initiated from seedlings germinated in vitro; they were identified by the abbreviation SS, for Sequoia seedling. The adult stocks were established by culturing shoot tips excised from trees that were at least 60 years
old. One stock, AS, was initiated from shoots isolated in 1976, and another, AST, was started from shoots obtained in 1994. Stocks of rejuvenated shoots, RS and 5XRT, were derived from AS and AST, respectively. The rejuvenation was achieved by repeatedly grafting the shoot tips from adult stocks onto freshly rooted segments of SS shoots in vitro. Five successive grafts resulted in the re-appearance of selected juvenile morphogenetic characteristics, namely high rooting competence and rapid shoot elongation, in the RS and 5XRT shoots. The grafting procedure, previously described by Huang et al. (1992), consisted of transferring ca. 1.5 cm long adult shoot terminals onto 1 cm long, rooted and decapitated juvenile shoots. Scions were left on rootstocks after each graft for 2 months. The new growths that emerged were then severed and their shoot tips re-grafted onto fresh rootstocks.
For ethylene analysis, 2 cm long stem terminals were subcultured in 25- × 150-mm glass tubes, each containing 20 ml nutrient medium. The medium contained MS (Murashige and Skoog, 1962) salts, 3% sucrose, 0.2% GelriteTM, and in µM: 555 i-inositol, 3 thiamine HCl, 2.4 each of nicotinic acid and pyridoxine HCl, and 26.6 glycine. After adjusting its pH to 5.7, diluting to volume, and adding and dissolving the Gelrite, prescribed quantities of medium were dispensed into culture tubes. Tubes were capped with Bellco polypropylene closures and autoclaved 10 min at 1.05 kg cm-2. The closures were not tight-fitting, so they enabled some gas exchange between the interior and exterior of tubes.
Experiments were performed at least 5 times. Ten cultures were employed for each of SS, AS, RS, AST, and 5XRT, with each culture containing one shoot. Three days prior to obtaining gas samples, the propylene closures were replaced by air-tight, serum vial caps to cause accu
1Corresponding author. Tel: (02) 2789-9590 ext. 324; Fax: (02) 2782-1854; E-mail: bolch@ccvax.sinica.edu.tw