Bot. Bull. Acad. Sin. (2000) 41: 267-273
Hsu et al. Effects of waterlogging on sudangrass
Effects of waterlogging on seed germination, electric
conductivity of seed leakage and developments of
hypocotyl and radicle in sudangrass
Fu-Hsing Hsu, Jeng-Bin Lin, and Shyh-Rong Chang
Department of Forage Crops, Taiwan Livestock Research Institute, Council of Agriculture, Hsinhua, Tainan, Taiwan, Republic of China
(Received October 15, 1999; Accepted March 13, 2000)
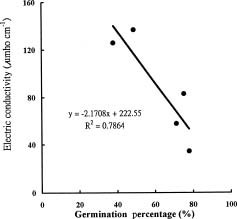
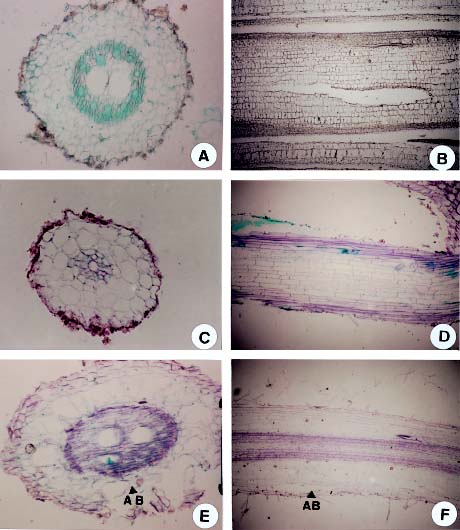
Abstract. Sudangrass (Sorghum sudanense Stapf) is a short-period forage species adapted to growth in Taiwan. The objectives of the experiment were to determine the effects of waterlogging on seed germination, electric conductivity of seed leakage and developments of hypocotyl and radicle in sudangrass and provide information for selecting the lines with tolerance to waterlogging stress. Sudangrass, CV. Taishi No. 1, was used for the experiment. In the germination test, the seeds were waterlogged for 3 days at 0, 1, 2 and 3 days after imbibing. The seeds waterlogged at 0 day after imbibing had the lowest germination percentage. The potassium contents of the seed leakages were increased and contents of both total sugar and reducing sugar in the seed leakages were decreased with an increase in imbibing period. The relationship between germination percentage and electric conductivity was negatively correlated (p<0.05), indicating that the electric conductivity could be used to evaluate the germination ability of sudangrass. For the emergence test, the sudangrass seeds were waterlogged for 0, 1, 2, 3, 4, 5, 6 and 7 days after sowing. The emergence percentages of sudangrass were not significantly different among treatments, while the emergence rate indices and the corrected emergence rate indices decreased with the increase in waterlogging period. This indicated that the emergence ability of sudangrass was reduced by waterlogging after sowing. The hypocotyl or the radicle cortex of sudangrass with a low germination or emergence percentage was damaged during waterlogging. The results demonstrated that: (1) the germination percentage of sudangrass was seriously affected when seeds were waterlogged before imbibing, (2) the electric conductivity of the seed leakage could be used to evaluate the germination ability of sudangrass.
Keywords: Electric conductivity; Hypocotyl; Radicle; Seed germination; Sorghum sudanense; Waterlogging.
Introduction
Sudangrass (Sorghum sudanense Stapf) is a short period forage species with high forage yield, drought tolerance, tillering ability, and fast regrowth. It has high amounts of crude protein and low amounts of crude fiber and hydrocyanic acid. It is adapted to growth in tropic and subtropic areas. Sorghum spp. has been used for forage in countries with a developed animal industry for a long time (McDonald et al., 1968; Sotomayor-Rios and Torres-Cardona, 1984).
The germination in this species is very sensitive to waterlogging. Decreases in germination ability have been attributed to a shortage of oxygen due to waterlogging (Orchard and Jessop, 1984). Respiration and electron transport are inhibited and ATP formation is decreased during germination when oxygen is deficient (Al-Ani et al., 1985; Johnson et al., 1989; Tsai et al., 1997). When the formation of ATP is reduced, the oxidation-reduction state between cell membranes becomes unbalanced and membrane permeability is increased. Thus, the solute
leakage is increased (Johnson et al., 1989), and electric conductivity is increased (Givelberg et al., 1984; Lott et al., 1991). The cell membrane deteriorates and the contents of carbohydrates, amino acids, organic acids and ions are increased in the leakage leading to an increase in electric conductivity when the seeds are waterlogged (Loomis and Smith, 1980; Mckersie and Stinson, 1980; Mukhear and Laidman, 1982; Simon and Raja-Harum, 1972). The seed vigor can be evaluated by the electric conductivity of the leakage (Agrawal, 1977; Yaklich and Abdul-Baki, 1975).
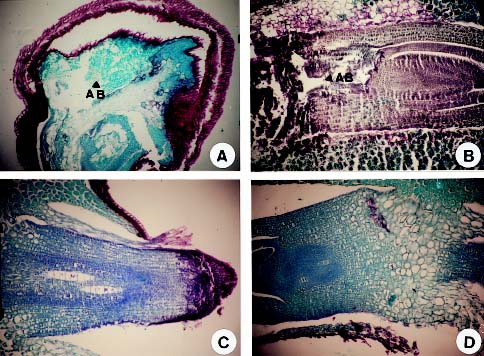
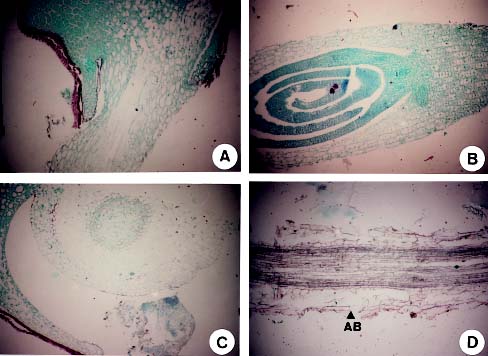
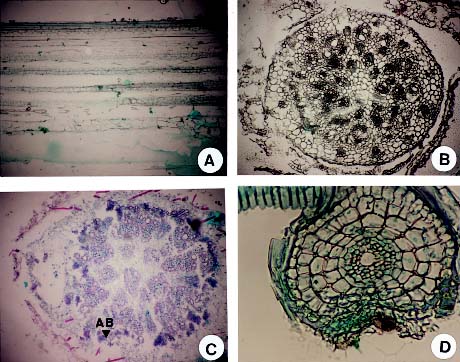
The root injury caused by waterlogging is due to oxygen stress in saturated soils (Drew, 1983). Drew et al. (1985) reported that many adventitious roots were formed when corn was under oxygen stress. In addition, air space development has been reported in the cortex of the root in several mesophytic species (Pezeshki, 1994). The objectives of this experiment were to determine the effects of waterlogging on seed germination, electric conductivity of seed leakage, and developments of hypocotyl and radicle in sudangrass. In addition, the results provide information necessary to develop screening tools for selecting the lines with tolerance to waterlogging stress, thus, improving field emergence and survival percentage of sudangrass during waterlogging.
1Corresponding author. Tel: (06) 5911211~250; Fax: (06) 5911182; E-mail: fhhsu@mail.tlri.gov.tw