Bot. Bull. Acad. Sin. (2000) 41: 283-291
Kuo-Huang et al. Sclereids in Nymphoides and Nuphar
Ultrastructural study on the formation of sclereids in the
floating leaves of Nymphoides coreana and Nuphar schimadai
Ling-Long Kuo-Huang1,2, Su-Hwa Chen1, and Shiang-Jiuun Chen1
1 Department of Botany, National Taiwan University, Taipei, Taiwan, Republic of China
(Received December 29, 1999; Accepted April 14, 2000)
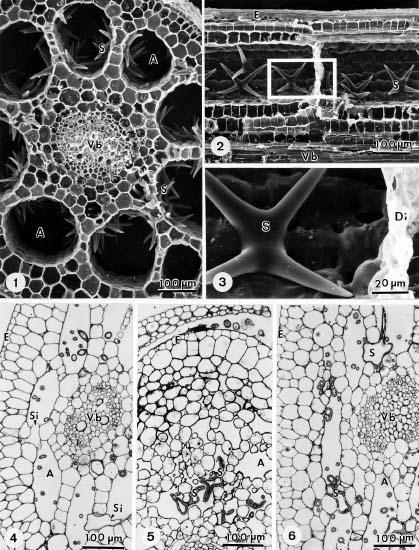
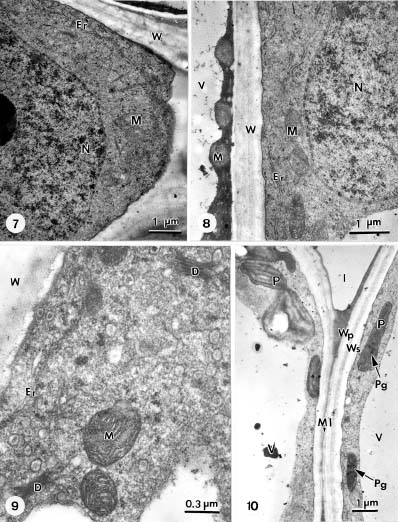
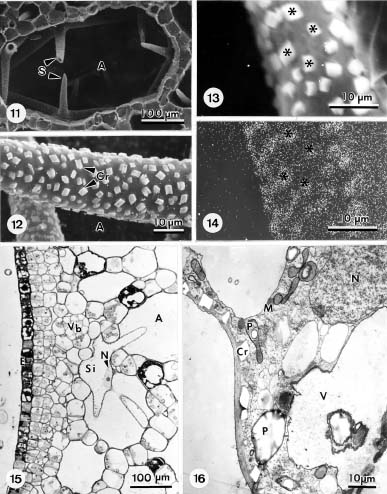
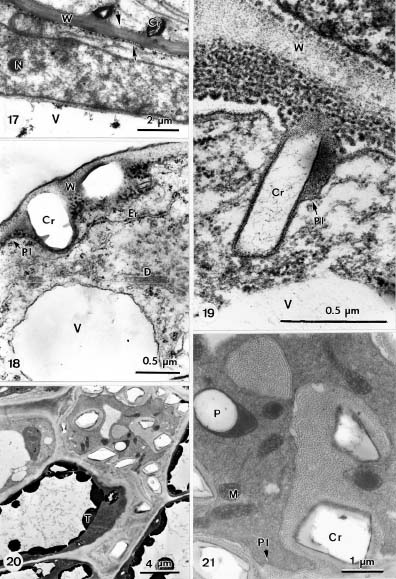
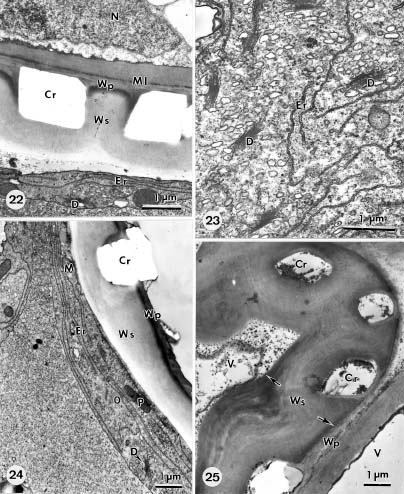
Abstract. The formation of star-shaped sclereids in the floating leaves of Nymphoides coreana and Nuphar schimadai was studied microscopically. These foliar sclereids were associated with the aerenchyma and found as the form of idioblast. The outer surface of mature sclereids was smooth in Nymphoides, but with many prismatic calcium oxalate crystals in Nuphar. However, the early morphogenesis of these two kinds of sclereids was similar. The sclereid initials were distinguished from the neighboring cells by their distinctly large nucleus. The expanding sclereid initials were constrained by the neighboring cells. Crystal formation in young sclereids of Nuphar started near the cessation of sclereid expansion. The crystals were bounded by crystal sheath and located in crystal chambers between the primary cell wall and plasma membrane. Calcium antimonate precipitates were found, especially on the crystal sheaths as well as between the plasma membrane and the primary cell walls. The crystal chambers have a paracrystalline appearance connected with the crystal sheath and the plasma membrane. After formation of crystals, the secondary wall was deposited and then the crystals became embedded between the primary and secondary walls. The possible functions of the foliage sclereids and the plans for further investigation are discussed.
Keywords: Calcium antimonate precipitates; Calcium oxalate crystals; Cell wall; Foliar sclereids; Nuphar schimadai; Nymphoides coreana.
Introduction
Sclereids can be found in many different parts of plant tissues and organs. They may serve various protective, strengthening, and other functions. For example, sclereids may form bands around seeds or cover roots or stems, possibly for strengthening purposes (Harris, 1983). Individual sclereids, called idioblasts, may cause tissues to be coarse and gritty and less palatable to insects (Mouseth, 1988). Such sclereids are frequently observed in the mesophyll of leaves of many plant genera, e.g., Borinia (Foster, 1955), Nymphaea (Gaudet, 1960; Kuo-Huang, 1992), and others (Rao and Das, 1979). The foliar sclereids of Olea europaea may also function as optical fibers to improve the light microenvironment within the mesophyll (Karabourniotis et al., 1994). The formation of sclereids can be observed during normal development of tissues and organs. However, differentiation of sclereids also can be induced artificially. For example, sclereids differentiated in the pith of Arabidopsis thaliana following repeated cutting of developing inflorescences (Lev Yadun, 1994), and a continuous pericyclic sclereid band developed in regenerated bark of Cinnamomum cassia, three years after the bark was removed (Hong and Tong,
1997). Various types of sclereids are found among different species, and sclereids in different tissues and organs have been used for plant classification (Rao and Banerjee, 1979; Villaron-Franceschinelli and Yamamoto, 1993).
The occurrence of sclereids in vascular plants has been studied mostly in terrestrial species, and rarely in aquatic species (Chiang and Huang, 1984; Kuo-Huang, 1990). In a preliminary study, we found many sclereids in the petiolated leaves of Nymphoides coreana (Menyanthaceae) and Nuphar schimadai (Nymphaeaceae). Both are members of the nymphaeid macrophytes (Smits et al., 1989). When comparing the foliar sclereids of these two aquatic species we found that their outer surfaces were quite different, which compelled us to investigate further. Results of light and electron microscopy are presented in this paper.
Materials and Methods
The Nymphoides coreana (L.) Hara (Menyanthaceae) and Nuphar schimadai Hayata (Nymphaeaceae) plants were grown in ponds located on the campus of National Taiwan University. Floating leaves in various stages were collected, and their petioles and laminae were cut into small pieces. The specimens were routinely processed through fixation (2.5-5% glutaraldehyde and 3-4% paraformaldehyde in 0.1 M sodium cacodylate buffer),
2 Corresponding author. Tel: (02) 2363-0231 ext. 2364; Fax: (02) 2391-8940; E-mail: linglong@ccms.ntu.edu.tw